Dopo i protobionti, le strutture organizzate - coacervati, microsfere, argille o altro - che erano riuscite a controllare in modo autonomo il bilancio energetico, che avevano acquisito un apparato genetico e riuscivano ad accrescersi e a dividersi, vennero selezionati positivamente fino ad avere delle protocellule. Queste protocellule dovevano essere molto diverse dagli attuali Procarioti, con un sistema genetico non ancora perfezionato e l'attività enzimatica ancora primitiva. Esse probabilmente erano eterotrofe, cioè assumevano le sostanze direttamente dal brodo primordiale; almeno questa è una delle ipotesi plausibili.
Le prime protocellule si estinsero e rimasero solo quelle che diedero origine all'ultimo antenato comune: L.U.C.A. (Last Universal Cellular Ancestor). Questo non significa che fosse il primo organismo esistito o che fosse l'unico, ma quello (o una comunità) che ha lasciato la sua eredità genetica oltre il Paleoarcheano (vedi carta stratigrafica).
{kind=link}
Le cellule rimaste, attraverso le mutazioni e la selezione naturale, affinarono l'apparato genetico, la membrana assunse una struttura più complessa, si aggiunsero nuove reazioni enzimatiche più specializzate fino ad arrivare alla cellula procariote.

Testimonianze fossili
Di queste primitive forme viventi non abbiamo alcuna traccia fossile. Gli sconvolgimenti geologici, l'attività esogena ed endogena, le radiazioni, hanno cancellato ogni possibile traccia fossile e, inoltre, non possedevano strutture atte alla fossilizzazione.
Le più antiche biotracce indirette sono state trovate in Groenlandia e hanno un'età di 3,83 miliardi di anni. Si tratta di particelle di grafite che potrebbero essere il prodotto di un'attività biologica di microrganismi che sfruttavano la CO2 atmosferica. È un reperto assai controverso, dunque, come lo è quello trovato in sedimenti rocciosi del Quebec, in Canada. In questo caso abbiamo microscopiche strutture tubulari di ematite, datate 3,77 - 4 miliardi di anni, simili a quelle si trovano nei pressi delle sorgenti idrotermali, lasciate da batteri più recenti.
I primi veri fossili sono delle strutture sedimentarie di acque poco profonde, chiamate stromatoliti, prodotte da batteri filamentosi, simili agli attuali cianobatteri.

Stromatoliti in Australia
Sono state trovate soprattutto in Australia occidentale e in Sudafrica e hanno un'età di 3,5 miliardi di anni. Si presentano con una successione verticale di lamine ondulate sovrapposte, la cui struttura è legata alla profondità dell'acqua.

Sequenza schematica della struttura interna di una stromatolite in base alla profondità dell'acqua (Crediti: clicca sull'immagine)
Ciascuna lamina è formata da uno strato di batteri e cianobatteri che intrappolano periodicamente fini sedimenti carbonatici, grazie alle mucillagini da essi prodotte.
Terminato uno strato, sopra la fanghiglia carbonatica si sviluppano altri filamenti che creano una nuova lamella.

All'epoca delle stromatoliti la vita era già ben affermata, perciò LUCA, l'ipotetico antenato comune a tutti i viventi, deve essere comparso prima di tale data, in un periodo compreso tra 3,6 e 4,1 miliardi di anni fa.
Eterotrofi o autotrofi?
Si suppone che i primi organismi fossero eterotrofi, anaerobi obbligati perché ancora non c'era l'ossigeno nell'atmosfera. Essi prelevavano il nutrimento dall'ambiente e non richiedevano meccanismi di biosintesi per produrre quanto necessario.
I Procarioti anaerobi impiegavano (e ancora lo fanno) processi di fermentazione, cioè degradazione incompleta degli alimenti, ricavando una piccola quantità di energia, però sufficiente per le loro attività metaboliche.
Con il trascorrere del tempo, però, il brodo diventava sempre più diluito e la produzione di nuove molecole organiche diminuì per l'affievolirsi delle scariche elettriche e delle altre fonti energetiche, perciò era necessaria una nuova via metabolica per rifornirsi di materia ed energia, quindi sopravvissero quelli che disponevano di una fonte di energia alternativa.
Altri autori, invece, sostengono che fossero autotrofe le prime forme viventi. Esse si sarebbero sviluppate intorno ai camini vulcanici sottomarini, sfruttando minerali come la pirite, acido solfidrico o idrogeno gassoso per fissare la CO2 e generare i composti organici. Questa opzione, tuttavia, è meno economica metabolicamente rispetto alla precedente e richiede reazioni molto sofisticate.
Queste due forme alternative non si escludono a vicenda. Le differenze che si osservano attualmente all'interno degli Archei e dei Batteri potrebbero essere spiegate proprio dallo sviluppo della vita nei pressi delle sorgenti idrotermali profonde e contemporaneamente nelle acque superficiali.
La fotosintesi
Per continuare a vivere in un ambiente che sta esaurendo i nutrienti, occorre una nuova via metabolica in grado di produrre sostanza organica catturando la luce solare: nasce la fotosintesi clorofilliana.
Si tratta di un passo evolutivo straordinario, avvenuto circa 3,5 miliardi di anni fa ad opera di cianobatteri (o batteri simili più antichi), che ha modificato radicalmente l'ambiente e i futuri organismi con l'immissione di ossigeno nell'atmosfera.
È anche un processo complesso che deve essere avvenuto in modo graduale, partendo da idrogeno atmosferico per fissare l'anidride carbonica (fotosintesi anossigenica) abbondante nella prima atmosfera (Batteri purpurei), per passare all'acido solfidrico (Batteri sulfurei), quando l'atmosfera si è modificata:
6CO2 + 12H2S + energia → C6H12O6 + 6H2O + 12S
Infine si giunse alla fotosintesi ossigenica, ricavando l'idrogeno direttamente dall'acqua (Cianobatteri).
6CO2 + 6H2O + energia → C6H12O6 + 6O2
La fermentazione deve avere accumulato quantità sempre maggiori di anidride carbonica, predisponendo le basi per il processo fotosintetico.
All'inizio, l'ossigeno, prodotto di scarto della fissazione dell'anidride carbonica liberata dalla fermentazione, ossidò il ferro presenta nell'acqua di mare, trasformandolo in ossidi insolubili che precipitarono, andando a formare depositi sui fondali e rocce rosse, schiarendo le acque che assunsero l'attuale colorazione.
L'ossigeno reagì successivamente anche con le rocce sedimentarie delle terre emerse contenenti ferro, formando delle stratificazioni a bande ferrose.

Formazione a bande di ferro nel Karijini National Park, in Australia
Dopo questo evento di ossidazione, avvenuto tra 2,45 e 1,850 miliardi di anni fa (Grande Ossidazione), l'ossigeno cominciò ad accumularsi nell'atmosfera, fino ad aumentare di 10.000 volte e raggiungere poi la concentrazione attuale a partire da circa 850.000 anni fa.

Come conseguenza di questo accumulo si ebbe l'ossidazione del metano atmosferico in anidride carbonica, che ha un effetto serra inferiore, raffreddando il pianeta fino al avere la grande glaciazione uroniana (Terra a palla di neve).
L'ossigeno, quindi fu prodotto inizialmente e per molto tempo dai Procarioti e solo in seguito dagli Eucarioti.
Per un periodo lunghissimo la vita continuò a prosperare negli oceani, dove l'acqua forniva una barriera protettiva contro i pericolosi raggi ultravioletti. Solo quando si formò uno strato di ozono, tra 1,850 e 0,850 miliardi di anni fa, i viventi poterono colonizzare anche le terre emerse.
Indipendentemente dal fatto che siano comparsi prima gli autotrofi o gli eterotrofi, la fotosintesi assicurò una grande quantità di nutrimento a tutti gli organismi viventi e senza di essa la vita si sarebbe estinta per mancanza di materie prime.
La respirazione cellulare
La Grande Ossidazione è detta anche Catastrofe dell'Ossigeno perché la tossicità dell'ossigeno causò una grande estinzione di massa delle prime forme di vita anaerobica.
La modifica della composizione atmosferica portò alla nascita di Batteri aerobi che sfruttavano l'ossigeno, attraverso la respirazione cellulare, per ricavare energia dagli alimenti in quantità molto superiore rispetto ai processi di fermentazione.
C6H12O6 + 6O2 → 6CO2 + 6H2O + energia
Abbiamo a questo punto dell'evoluzione:
- gli autotrofi che si nutrono autonomamente sfruttando l'energia solare (organismi fotosintetici, oppure quella ricavata da sostanze inorganiche (chemiosintetici);
- gli eterotrofi che dalle sostanze organiche ricavano energia attraverso la respirazione, ma rimangono ancora alcuni organismi che si servono della fermentazione.
La cellula eucariote
Una nuova tappa evolutiva ha portato, circa 1,5 - 1,4 miliardi di anni fa, alla cellula eucariote.
Attraverso una fase autogena si è formato il nucleo e gli altri organelli dotati di membrana, come il reticolo endoplasmatico e le altre vescicole, mediante introflessioni della membrana plasmatica, nel cui spessore sono inclusi enzimi e altre molecole partecipanti al metabolismo.

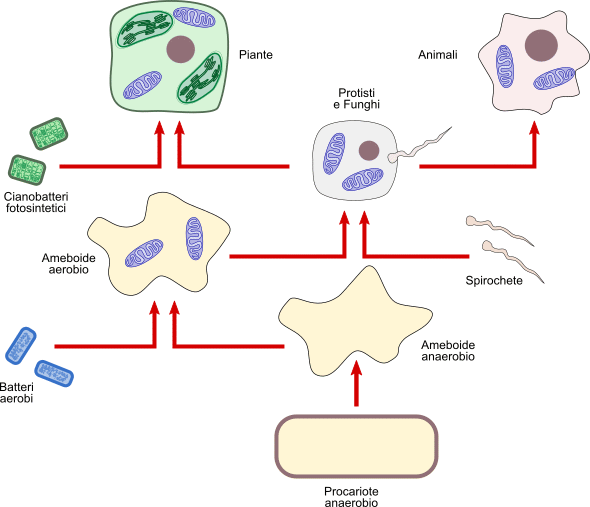
Per quanto riguarda i mitocondri e i cloroplasti, invece, la biologa statunitense Lynn Margulis (1938 - 2011) ha pubblicato nel 1970 il libro Origin of Eukaryotic Cells, in cui enunciava la teoria endosimbiontica seriale.
Descrivendo questi due organelli, avevamo visto come essi possedessero un proprio DNA di tipo procariote e si duplicassero in modo autonomo. Secondo questa teoria, si suppone che originariamente fossero dei Procarioti autonomi che sono stati inglobati da un'altra cellula.
La prima fase è consistita nella perdita della parete cellulare di una grande cellula procariote anaerobia (Fagocita primitivo) la cui capacità fermentativa forse non era più in grado di soddisfare le richieste di energia e alla quale l'ossigeno, sempre più presente nell'atmosfera, stava diventando tossico.
La perdita della parete ha consentito l'ingresso di una Battere aerobio più piccolo, avente la capacità di utilizzare l'ossigeno per demolire le sostanze organiche (in particolare il glucosio) attraverso la respirazione cellulare. Questo primo inglobamento può essere avvenuto per un atto di parassitosi oppure di fagocitosi da parte della cellula ospite. In entrambi i casi, il Battere per qualche motivo non è stato digerito ed è iniziata una collaborazioni di simbiosi in cui il Fagocita riceveva l'energia in eccesso della respirazione batterica e si liberava del tossico ossigeno, aumentando la capacità di sopravvivenza, mentre il Battere veniva a trovarsi in un ambiente protetto, non aveva più la necessità di andare alla ricerca di cibo e poteva delegare alcune funzioni metaboliche alla cellula ospite e anche parte delle informazioni genetiche: nasce così il precursore del mitocondrio. Attraverso questa tappa si arriva alla cellula eucariote dei Funghi e degli Animali.
Un secondo inglobamento riguarda la simbiosi della nuova cellula eucariote con Batteri dotati di mobilità, simili alle attuali Spirochete. Con questa simbiosi, alcune cellule eucariote acquisirono la capacità di spostamento, utile per la ricerca del cibo. Si formarono, infatti, ciglia e flagelli, dotati di proteine contrattili, tipici di alcuni Protisti e alghe, centrioli e microtubuli e anche alcuni tessuti di Metazoi. Interessante il fatto che i centrioli siano in grado di duplicarsi autonomamente e perciò potrebbero contenere piccole quantità di DNA (non confermata la sua scoperta nel 1990) a riprova della loro origine autonoma.
Una terza incorporazione - per questo si parla di endosimbiosi seriale - ha coinvolto un Procariote fotosintetico (Cianobattere), dando origine al cloroplasto, organello tipico delle Piante.
Queste simbiosi è probabile che all'inizio non fossero permanenti e ci sono voluti 2 - 3 miliardi di anni per arrivare alla cellula eucariote come oggi la conosciamo, dotata dei suoi numerosi organelli.
Dopo l'esposizione della teoria dell'endosimbiosi, possiamo vedere come sono oggi suddivisi sistematicamente gli organismi viventi. La classificazione può variare tra i diversi autori.
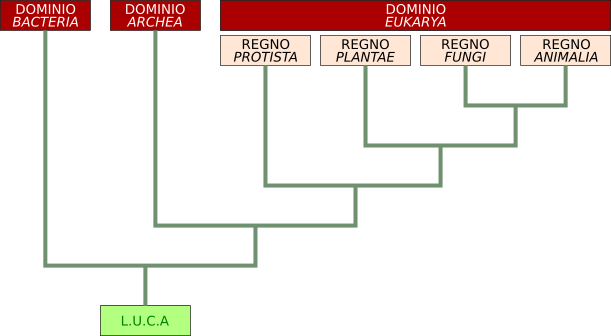
Una prima classificazione divide i viventi in due grandi domini: Prokaryota ed Eukarya.
Il primo comprende i 2 regni Bacteria e Archea (alcuni lo indicano come unico regno delle Monere), mentre il secondo comprende 4 regni: Protista, Plantae, Fungi, Animalia.
Considerando però le relazioni evolutive, altri autori individuano una prima linea che ha portato ai Bacteria, elevati al rango di dominio, e l'altra all'antenato comune di Archea (anche questi elevati a dominio) ed Eukarya (disegno sotto). Gli Archei, infatti, pur essendo Procarioti, sono più vicini alle cellule eucariote e per questo sono stati inseriti in un dominio separato.
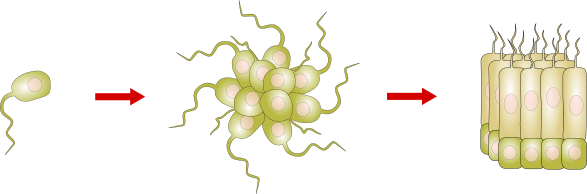
La pluricellularità
Diversamente dalle fasi evolutive precedenti, il passaggio ad organismi pluricellulari deve essere stato abbastanza rapido, solo qualche centinaio di milioni di anni. Infatti, 670 milioni di anni fa a Ediacara (Australia) era ben rappresentata una ricca fauna e nel Cambriano erano presenti i principali phyla di animali e piante. La più antica prova fossile di pluricellularità è datata a circa 1,2 miliardi di anni. Si tratta di un'alga rossa, la Bangiomorpha pubescens, rinvenuta in Canada.
Per essere considerato pluricellulare, un organismo deve avere più cellule unite tra loro, che comunicano, che dipendono l'una dall'altra e che siano differenziate.
La pluricellularità non è stata acquisita una sola volta, ma si è evoluta indipendentemente almeno una ventina di volte nei viventi e, probabilmente, con modalità differenti. Si possono comunque individuare due modalità principali.
Un primo modo è quello della divisione e adesione: una cellula si divide in due celle figlie mediante mitosi che poi rimangono unite dalla stessa membrana. Ciascuna di esse può successivamente specializzarsi a compiere funzioni diverse, mantenendo una stretta collaborazione e comunicazione. Oppure, un Protista di grandi dimensioni e plurinucleato sviluppa al suo interno delle pareti che delimitano i diversi nuclei e ciascuno diventa una cellula indipendente.

Un secondo modo è quello dell'aggregazione. Un gruppo di Protisti unicellulari si aggrega per formare una colonia. Quando le cellule si dividono, i discendenti rimangono attaccati e nella colonia alcune cellule si specializzano, acquisendo specifiche funzioni.