
Ogni teoria è valida se presenta una serie di prove a suo sostegno. La teoria dell'evoluzione ha a suo favore un'ampia serie di prove, anche se, come abbiamo detto all'inizio della sezione, i creazionisti non le ritengono sufficienti o valide.
I fossili
I fossili sono i resti di organismi del passato che noi rinveniamo all'interno degli strati rocciosi.
La fossilizzazione è un processo eccezionale: solo un numero infinitesimo degli organismi fossilizza e di ciascuno rimangono le parti più dure, come le ossa e i gusci. La conservazione è favorita in ambiente marino mentre in quello terrestre, se non viene immediatamente sepolto, l'organismo è distrutto dai predatori, dai decompositori e dagli agenti atmosferici. Nel corso del tempo si sono succeduti miliardi e miliardi di organismi: se non venissero decomposti, nel giro di pochissimo tempo non avremmo sostanza organica sufficiente per generare nuovi organismi.
Nonostante la rarità delle condizioni idonee alla fossilizzazione, sono state trovate circa 250000 specie fossili, molte delle quali con milioni di esemplari, che rappresentano tuttavia l'1% delle specie esistite.
Darwin ha fatto dei fossili uno dei punti fondamentali della sua teoria, ma si lamentava della loro scarsità, esponendo le possibili cause; ne aggiungiamo qualcuna: fenomeni erosivi e tettonici, organismi a corpo molle, le popolazioni in cui si verifica un fenomeno evolutivo sono generalmente piccole e quindi a bassa probabilità di fossilizzazione.
Nel corso di decenni gli scaffali dei musei e dei dipartimenti si sono enormemente arricchiti di esemplari fossili, per cui oggi abbiamo una buona sequenza temporale e spaziale della distribuzione delle specie.
Esaminando i reperti trovati negli strati geologici, possiamo stabilirne sia una datazione relativa, in base alla successione degli strati, sia assoluta, con il metodo radiometrico. Nella sequenza stratigrafica si nota che negli strati più antichi si trovano pochi fossili di organismi marini semplici, mentre quelli soprastanti contengono organismi più complessi, in maggiore numero e varietà; in quelli più recenti le forme sono simili alle attuali Questa successione è spiegabile con la progressiva trasformazione e complessificazione delle forme.
Se consideriamo i Vertebrati, vediamo che si sono evoluti nell'ordine: i Pesci, gli Anfibi, i Rettili e poi Uccelli e Mammiferi. Anche per le Piante si trovano prima i Muschi e poi le Angiosperme. La progressione evolutiva sarebbe sconfessata se venissero trovati organismi complessi come i Primati accanto ai Dinosauri, oppure Mammiferi prima dei Pesci, ma finora non è mai accaduto. Sono comunque poche le forme che si sono evolute verso una maggiore complessità: la moda, cioè il valore più frequente, rimane sempre fissa su Batteri.
Dal punto di vista temporale, abbiamo già visto nelle pagine precedenti in una stessa area si possono rinvenire fossili simili alle forme moderne. È il caso del Gliptodonte, unico mammifero corazzato, trovato da Darwin in Sudamerica, che ha un corrispondente nel più piccolo Armadillo (Dasypus novemcinctus), presente oggi nei medesimi luoghi.


Ricostruzione di
Doedicurus
e
Glyptodon
- Moderno Armadillo
La successione di specie affini nel corso del tempo si spiega con le continue variazioni ambientali avvenute nel corso della storia della Terra, per cui si ha un progressivo adattamento delle specie con il meccanismo della selezione naturale, mentre altre si sono estinte.
Lo studio dei fossili permette di stabilire legami di parentela tra specie vissute in passato e quelle attuali. Le differenze fra queste diventano maggiori più si retrocede nel tempo secondo una serie graduale, ma a volte repentina, di cambiamenti delle caratteristiche anatomiche che li collegano tra loro: ciò dimostra la divergenza da antenati comuni.
Per alcuni gruppi di animali i fossili sono abbondanti e ben distribuiti per cui si possono ricostruire le linee evolutive: ne sono un esempio l'evoluzione degli Equidi, dei Camelidi e dei Proboscidati in cui è possibile ricostruire l'intera sequenza di adattamenti che hanno portato forme ancestrali a evolversi nelle forme attuali, evidenziando caratteristiche intermedie tra le specie attualmente viventi e gli organismi del passato. Va però fatto presente che alcuni autori criticano fortemente tali sequenze ricostruite.

Il processo complesso di evoluzione del cavallo, che ha portato a un aumento di statura, lo sviluppo del terzo dito e la modifica della dentatura.
Molto più numerose sono le linee evolutive degli Invertebrati marini, dove la fossilizzazione è molto più facile, come nel caso dei Trilobiti, delle Ammoniti e di molti microfossili.
Non sempre è possibile trovare le forme intermedie tra due specie, come già aveva evidenziato Darwin e quindi si parla di anelli mancanti. Quando si vuole risalire a un antenato di due specie divergenti, ad esempio uomo e scimpanzé, non si devono cercare le forme intermedie tra queste due specie, che non esistono perché l'una non deriva dall'altra, ma la specie antecedente che presenta caratteristiche comuni a entrambe. Utilizzando l'esempio che fece Darwin, se considero il tapiro e il cavallo, non devo cercare le forme intermedie tra questi due, ma gli intermedi tra il tapiro e un antenato comune e tra il cavallo e lo stesso antenato e questo avrà caratteristiche sia del tapiro che del cavallo.
In alcuni casi gli anelli sono effettivamente mancanti e questo può dipendere da due cause.
- L'anello non è stato trovato perché esistono ancora moltissime zone non esplorate e, come dice Darwin, «l'assenza di una prova non è la prova della sua assenza».
- Gli anelli possono non esistere perché, secondo il modello evolutivo degli equilibri intermittenti, la speciazione in alcuni casi è stata così rapida geologicamente da non aver consentito la fossilizzazione di forme di transizione.
Per quanto riguarda i Vertebrati, possediamo diversi anelli di congiunzione.
L'anello tra Pesci e Anfibi è rappresentato da Tiktaalik roseae, vissuto circa 375 milioni di anni fa e scoperto nel 2004. Questo animale aveva il capo appiattito come le salamandre, con narici e occhi sulla cima del capo, il collo (che non hanno i pesci), una fessura branchiale, forse anche polmoni primitivi, scaglie e pinne lobate proprie dei pesci. Le pinne anteriori, però, sono robuste e analoghe all'arto dei coccodrilli, perciò era in grado di sostenersi anche se non proprio di camminare.

(Cediti:
ponizej/CC BY-SA 3.0
)
{kind=link}
La successiva forma di transizione è Ichthyostega, che possiede arti meglio definiti rispetto al precedente, ma tozzi e brevi, assomiglia a una grossa salamandra.

Per il passaggio da Rettili a Uccelli abbiamo tre generi: Archaeopteryx, Microraptor, Sinornithosaurus.
Il famoso Archaeopteryx lithographica, trovato in Germania nel 1860, delle dimensioni di un corvo, ha caratteristiche da rettile per quanto riguarda lo scheletro (mascelle con denti, artigli, coda ossea, dita separate nelle ali), ma possedeva grandi penne e il primo dito delle zampe opponibile.


Microraptor gui, rinvenuto in Cina, aveva vere e proprie penne di colore nero con iridescenze, forse utili per il corteggiamento e la termoregolazione.

Nel 2010 è stato trovato un esemplare di Sinornithosaurus che aveva conservato perfettamente le piume provviste di rachide e anche il colore, che era rosso mattone con sfumature di giallo, grigio e nero. Probabilmente le usava per planare da un albero all'altro.

Le specie che testimoniano il passaggio da Rettili a Mammiferi appartengono ai Cinodonti, animali vissuti alla fine dell'era Paleozoica, comparsi prima dei Dinosauri.
Presentano molte caratteristiche dei Mammiferi, come il corpo ricoperto di pelo, una dentatura tipicamente carnivora, il palato separa la cavità boccale da quella nasale, probabilmente possedevano vibrisse; l'articolazione della mandibola con la mascella non è ben distinguibile ma, secondo la modalità di articolazione, vengono avvicinai ai Rettili o ai Mammiferi; le dimensioni variavano, secondo la specie, da quelle di un roditore a quelle di un cane; l'andatura era in alcuni casi simile a quella dei Rettili, con zampe ai lati del corpo, mentre altri si spostavano come i Mammiferi; erano probabilmente omeotermi e deponevano le uova come i Rettili.

Prove anatomiche e morfologiche
L'anatomia comparata è la disciplina che confronta le strutture anatomiche di diversi organismi.
La comparazione dei gruppi animali o vegetali implica la presenza di un piano comune di organizzazione del corpo e, nel caso delle omologie, la discendenza da un antenato comune.
Da questo piano strutturale di base la selezione naturale ha portato successivamente l'adattamento dell'organismo alle diverse funzioni, trasformando, sviluppando, riducendolo o facendo scomparire alcune parti del corpo.
Le omologie
Organi omologhi sono quelli che hanno la stessa struttura, avendo la medesima derivazione embrionale, ma funzione diversa.
Si possono identificare gli stessi organi in un gruppo sistematico, anche se sono modificati per volume, forma e funzioni in base alle condizioni di vita.
Se noi osserviamo, ad esempio, arti dei Vertebrati tetrapodi, cioè Anfibi, Rettili, Uccelli e Mammiferi, possiamo riconoscere sempre la medesima struttura di base, chiamata arto pentadattilo, costituita da tre segmenti, a loro volta suddivisi in un osso prossimale (omero), due ossa distali (radio e ulna), una serie di ossa carpali, cinque serie di ossa metacarpali e falangi. Questo conferma la derivazione da un antenato comune dal quale hanno ereditato tali strutture.
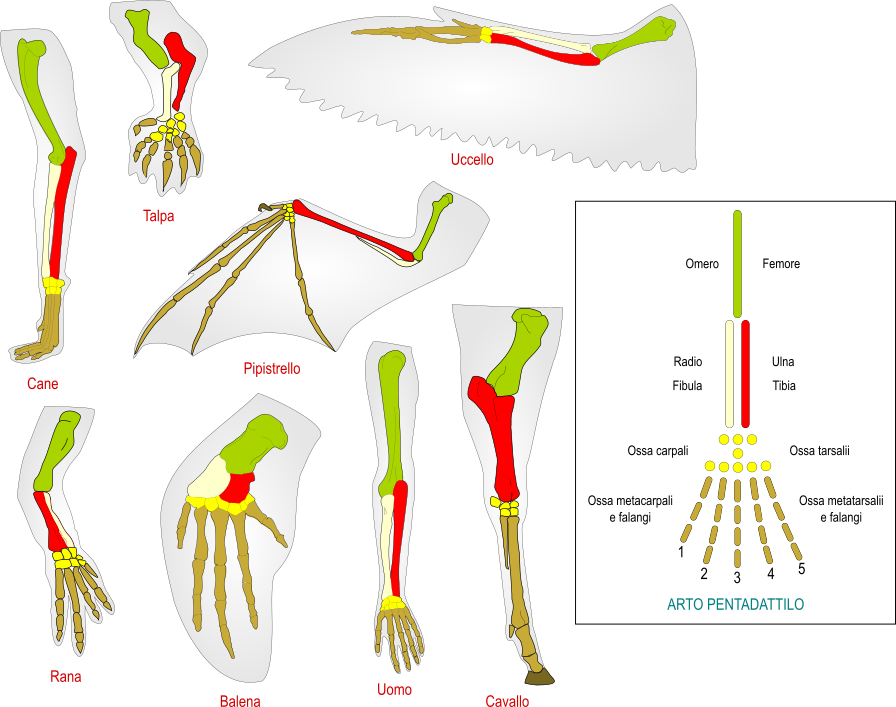
(Clicca sull'immagine per ingrandirla)
Nei Primati gli arti anteriori sono modificati per afferrare. Nel cane e nella rana gli arti anteriori e posteriori servono a deambulare. Nel cavallo si ha un allungamento del terzo dito e la presenza di uno zoccolo. Gli arti della talpa sono corti e adatti a scavare. Nei Cetacei gli arti sono modificati per nuotare. Nei pipistrelli quattro dita si sono allungate per sostenere il patagio, mentre il primo dito presenta un gancio per afferrare. Negli Uccelli gli arti anteriori si sono adattati per il volo.
Anche nei fiori troviamo sempre sepali, petali, ovario, stilo e stigma, anche se varia il numero, la forma e il colore di ciascun elemento secondo la specie.
Le analogie
Si definiscono analoghi gli organi aventi funzioni simili ma strutture anatomicamente diverse, poiché hanno un'origine embrionale differente, non appartenendo alla stessa linea evolutiva. Le analogie le ritroviamo in specie in cui la selezione ha favorito i medesimi adattamenti, in quanto vivono nello stesso tipo di ambiente.
Ad esempio, le ali degli Insetti e degli Uccelli assumono forma esteriore e funzioni analoghe, pur non derivando dagli stessi territori embrionali.

Organi vestigiali e atavismi
Gli organi rudimentali o vestigiali sono strutture in via di riduzione che hanno perso la loro funzione oppure svolgono una funzione diversa da quella originaria; sono propri di tutti gli individui della stessa specie e omologhi a strutture corrispondenti in gruppi affini.
Gli organi vestigiali sono abbastanza frequenti sia nelle piante sia negli animali. Vediamo alcuni esempi.
Le talpe hanno occhi ridotti e non funzionanti sia perché possono danneggiarsi, sia perché la loro formazione richiede un notevole dispendio energetico.

Gli animali che vivono nelle grotte come ragni, salamandre, gamberetti ecc. hanno occhi assai ridotti o del tutto assenti.
Le balene hanno cinto pelvico incorporato nei muscoli e arti posteriori ridotti, che dimostrano la discendenza da antenati terrestri.

I serpenti hanno femore e ossa del bacino, anche se sono privi di arti.
Le ali dello struzzo, emù, casuario, kiwi, hanno ali ridotte, non adatte per il volo in quanto questi animali non hanno molti predatori, un vantaggio evolutivo perché il volo è molto dispendioso.

Emù
Nelle piante si può presentare un androceo non funzionante nei fiori femminili o un gineceo in quelli maschili.
Nell'uomo abbiamo l'appendice ciecale molto ridotta, mentre quella omologa dei Mammiferi erbivori è assai sviluppata perché contiene i Batteri necessari alla digestione della cellulosa.
La plica semilunare è una membrana presente nell'occhio dei Mammiferi, residuo della membrana nittitante, mentre Negli Anfibi, nei Rettili e negli Uccelli ha l'importante funzione di protezione e idratazione.
Il tubercolo di Darwin è un ispessimento cartilagineo del padiglione auricolare, residuo di un'articolazione che permetteva ai progenitori di orientare le orecchie.
Alle orecchie sono attaccati tre muscoli che in passato consentivano il movimento delle orecchie.
Il coccige, ultimo segmento della colonna vertebrale, è formato da 4 - 6 abbozzi di vertebre saldate tra loro. È quanto rimane della coda presente nei nostri antenati.
Altre vestigia: mammelle nei maschi, muscoli del naso, muscoli erettori del pelo, pelo sul petto, denti del giudizio, segmentazione dei muscoli addominali.
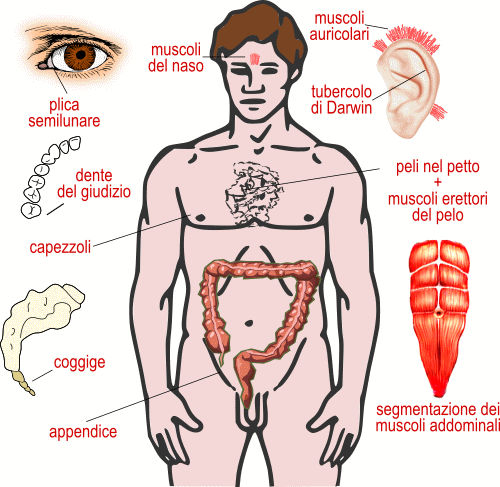
Organi vestigiali nell'uomo
Gli atavismi sono strutture presenti solo in alcuni individui di una specie. Si tratta di caratteri presenti negli antenati, che ora non si formano più. Occasionalmente essi ricompaiono perché i geni si sono conservati, anche se normalmente repressi.
Ad esempio, possono comparire arti posteriori nelle balene o in Vertebrati normalmente privi di arti. A volte si notano denti nei polli, speroni nei cani, dita aggiuntive nei cavalli.

Sperone nel cane
Negli embrioni è presente la coda, che scompare dopo sette settimane e le vertebre si fondono insieme nel coccige. Negli esseri umani può comparire una vera e propria coda, con vasi sanguigni nervi e muscoli che consentono il movimento. Possono comparire anche capezzoli soprannumerari.

L'embriologia
L'embriologia è la scienza che studia i processi di sviluppo degli embrioni. In particolare, l'embriologia comparata confronta lo sviluppo degli embrioni di diversi organismi.
La legge biogenetica fondamentale formulata da Ernst Haeckel (1834 - 1919) nel 1868 afferma che: «L'ontogenesi, o evoluzione individuale, è una breve e rapida ricapitolazione della filogenesi o dello sviluppo del gruppo corrispondente, vale a dire degli antenati dell'individuo, e questa ontogenesi si effettua conformemente alle leggi dell'eredità dell'adattamento».
Secondo questo zoologo, quindi, i processi di sviluppo (ontogenesi) di un organismo ricapitolano la storia evolutiva del gruppo a cui appartiene e questa sarebbe la prova di un antenato comune.
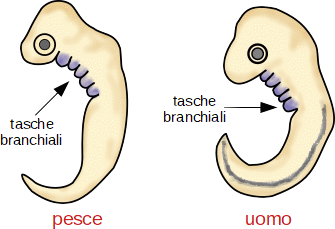
L'esempio classico è quello fornito dallo studio embriologico di Pesci, Anfibi, Rettili, Uccelli, Mammiferi. Se confrontiamo gli stadi precoci di queste cinque classi di Vertebrati, notiamo una forte somiglianza tanto da essere indistinguibile l'embrione di un Anfibio da quello di un Mammifero. In particolare, tutti gli embrioni presentano tasche branchiali e un abbozzo di coda.
Nel corso dello sviluppo le tasche branchiali si trasformano in modo di verso. Mentre nei Pesci si fessurano diventando branchie, nei Mammiferi si trasformano nella membrana del timpano, nella tromba di Eustachio, in alcune ossa e cartilagini della mandibola e nelle paratiroidi.
L'abbozzo della coda diventa una coda vera e propria in alcune specie, mentre in altre regredisce.

Disegni degli stadi di sviluppo embrionale erroneamente attribuiti a Haeckel
La teoria della ricapitolazione è oggi da considerarsi scientificamente falsificata.
Lo sviluppo embrionale di un organismo di una data specie non ripercorre in breve le tappe evolutive di quella specie: la formazione delle diverse strutture non avviene in modo lineare, ma segue vie contorte, tracce di antichi percorsi evolutivi assai più complicate di quelle proposte da Haeckel. I nuovi dati non avvallano la teoria, anzi, non è possibile scorgere un antenato in nessuno stadio dello sviluppo embrionale. Gli embrioni umani possono somigliare per certi versi a quelli dei pesci, ma nel corso del loro sviluppo non somigliano mai a uno dei nostri antenati.
L'embriologia invece dimostra l'esistenza di uno schema di base, a partire dal quale si formano strutture completamente diverse grazie alla presenza di specifici geni che controllano lo sviluppo. Nelle prime fasi entrano in azione i primi geni essenziali, ad esempio i geni Hox, poi si attivano via via gli altri geni specifici che determineranno il fenotipo definitivo.
La moderna biologia evolutiva dello sviluppo (evo-devo) ci mostra l'importanza che ha lo sviluppo embrionale nella modifica del fenotipo adulto, ma questo argomento lo affronteremo più avanti.
La Biologia molecolare
Le prove fornite dalla biologia molecolare sono le più recenti, ignote a Darwin, ma sicuramente le più importanti.
Gli organismi, nonostante la grande varietà morfologica, mostrano una notevole uniformità quando si scende a livello cellulare e, più ancora, a livello biochimico.
- Tutti gli esseri viventi sono essenzialmente composti da quattro elementi chimici (carbonio, ossigeno, idrogeno, azoto) ai quali si aggiungono altri elementi presenti in maggiore o minore quantità.
- Tutti sono costituiti da quattro tipi di macromolecole (protidi, lipidi, glucidi, acidi nucleici) le quali rappresentano la base di tutti i processi metabolici.
- Tutti hanno la medesima unità strutturale: la cellula costituisce la base comune della struttura dei viventi.
- In tutti gli organismi, il codice genetico è lo stesso (tranne rari casi). È dunque universale. Potrebbe funzionare ugualmente bene, anche se fosse diverso tra le specie, ma il fatto che sia uguale testimonia una parentela.
- Gli acidi nucleici hanno la medesima struttura in tutti gli organismi e anche il contenuto è simile, tanto che è possibile ibridare le molecole di specie diverse: quanto più è saldo l'appaiamento, tanto più le specie sono evolutivamente vicine.
- Lo stesso tipo di geni si trova in molti organismi, come i geni omeotici che controllano lo sviluppo e sono presenti grandi quantità di DNA non codificante, fra cui gli pseudogeni, cioè copie imperfette e lesionate che rappresentano vestigia evolutive di geni che hanno subito alterazioni mutazionali significative.
- C'è un'unità funzionale: gli organismi hanno i medesimi processi metabolici di base, come la glicolisi, la sintesi proteica, ecc.
- La fonte energetica di tutti gli organismi è l'ATP.
- Anche i retrovirus endogeni forniscono una prova dell'evoluzione. Nel genoma si trovano delle sequenze residue lasciate da antiche infezioni virali. Queste sequenze vengono sempre trasmesse alla generazione successiva di quell'organismo che ha ricevuto l'infezione. Poiché questo evento è raro e casuale, quando si trovano retrovirus in posizioni cromosomiche identiche in due specie diverse, ciò suggerisce un'origine comune.
- Le proteine di ogni vivente sono composte dagli stessi 20 amminoacidi, anche se teoricamente potrebbero essere di più e diversi tra le specie. Inoltre, sono tutti in forma L, pur essendo funzionalmente identici quelli in forma D. Probabilmente, le prime forme viventi avevano casualmente amminoacidi con la forma L, che è stata poi trasmessa alla discendenza.
- Alcune proteine sono talmente importanti da trovarsi pressoché in tutti gli organismi viventi. È il caso delle polimerasi, delle proteine dei ribosomi ecc. Si noti come al salire della complessità dell'organismo, non si modifica la struttura base di queste proteine, ma si aggiungono delle subunità. Le leggere differenze o le somiglianze consentono di stabilire le affinità tra le specie e i legami evolutivi.
L'osservazione comparata della struttura primaria di proteine appartenenti a organismi molto diversi ci indica quante mutazioni, cioè quante sostituzioni di amminoacidi, le separano. Ad esempio, il citocromo c, essenziale per la respirazione cellulare, è formato da circa 104 amminoacidi. Nel tempo, la proteina ha subito mutazioni nel tipo di amminoacidi senza alterarne la funzionalità (altrimenti l'organismo non sopravviverebbe): se noi confrontiamo il citocromo c di due specie, in base al numero di amminoacidi modificati è possibile stabilire da quanto tempo essi si sono diversificati: quanto maggiore è il numero di differenze, tanto più lontano è l'antenato comune da cui derivano. Ad esempio, l'uomo e lo scimpanzé hanno la proteina identica, mentre tra questi e la scimmia Rhesus (Macaca mulatta) la differenza è di un amminoacido. Tra il citocromo c del cavallo e quello di un insetto c'è un certo numero di differenze, ma sono molte di più tra il cavallo e il frumento.

Applicando questa metodologia a diversi tipi di proteine si arriva a datare il momento in cui da una specie ancestrale si sono staccate nuove specie. Le proteine si comportano pertanto come orologi molecolari: contando le sostituzioni in amminoacidi, si determina il tempo trascorso dal distacco dall'antenato comune, assumendo che le mutazioni si accumulino con velocità costante nel DNA.
Oggi il procedimento è semplificato dalle moderne tecniche di sequenziamento del DNA, che consentono di comparare interi genomi i cui risultati sono molto affidabili e confermano sempre più l'ipotesi di un'origine comune. Le specie più strettamente imparentate condividono molte più sequenze di quelle filogeneticamente più lontane.
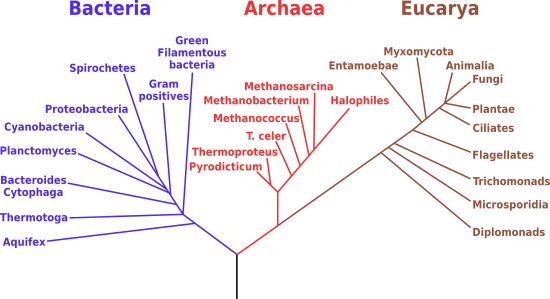
Attraverso il sequenziamento è possibile costruire un albero filogenetico, cioè il diagramma che mostra le relazioni fondamentali di discendenza comune di gruppi tassonomici di organismi (vedi immagine sopra e anche sotto).
Ogni albero ha una radice, che indica la popolazione da cui derivano le specie in esso rappresentate, cioè l'antenato comune; i rami rappresentano il percorso evolutivo di un determinato gruppo, specificando antenati e discendenti; gli apici sono le forme estinte o ancora viventi, a seconda del punto in cui arrivano; i nodi sono i punti di divergenza di due gruppi, distinti morfologicamente e geneticamente, che originano discendenti separati e rappresentano l'antenato comune.

L'albero filogenetico è generalmente congruente con quello della tradizionale tassonomia e paleontologia e, ove non lo sia, si provvede alla correzione in base ai nuovi dati genetici.
Biogeografia
La biogeografia è la disciplina che studia la distribuzione delle piante e degli animali nei diversi luoghi del mondo.
Abbiamo già trattato l'argomento nella pagina di Darwin, a cui si rimanda. Riprendiamo qui qualche concetto.
La distribuzione delle specie è assai singolare perché non troviamo le stesse forme di vita in tutti gli ambienti nei quali risulterebbero adattati, al contrario, le forme viventi occupano aree ben precise, spiegabili solo con la teoria evolutiva combinata con la Tettonica delle placche, a meno che non si voglia pensare ad un'assurda bizzarria del Creatore. Ogni gruppo si è evoluto in una determinata area e poi si è diffuso in luoghi lontani se si sono presentate possibilità di spostamento oppure sono rimasti confinati, in presenza di barriere geografiche. Se non ci fosse un collegamento tra la distribuzione delle specie e l'evoluzione, ci si aspetterebbe di trovare una determinata specie in ogni luogo dove si possa adattare e sopravvivere.
Ad esempio, le specie africane sono totalmente diverse da quelle del Sud America, eppure si trovano alla medesima latitudine. Specie fossili, invece, possono trovarsi anche su continenti diversi, questo perché in passato formavano un unico supercontinente come la Gondwana, poi separatasi per il movimento delle placche tettoniche (si vedano le prove della deriva dei continenti).
Il cactus e l'euforbia sono due specie analoghe come morfologia e occupano il medesimo tipo di ambiente, ma il primo si trova solo nei deserti nordamericani, mentre la seconda solo in quelli africani.
L'Australia è priva di Placentati, ma è l'unico continente ad avere le circa 60 specie di canguri e la gran parte dei Marsupiali.
La Nuova Zelanda e le isole oceaniche sono prive di Mammiferi terrestri e quando vi sono stati introdotti dall'uomo hanno prosperato fino a minacciare le specie endemiche.
L'orso bianco si trova esclusivamente attorno al Circolo Polare Artico e i pinguini solo al Polo Sud e non viceversa.

Prove fornite dagli esperimenti
Di solito si pensa che l'evoluzione sia lenta e graduale e che per questo non sia possibile osservarla direttamente. Anche se generalmente le trasformazioni avvengono nella scala dei tempi geologici, molte prove a favore dell'evoluzione sono state fornite in tempi recenti dai risultati di esperimenti e da osservazioni, usando organismi che hanno una riproduzione molto veloce.
Un esperimento su larga scala è quello del E. coli Long-Term Evolution Experiment (LTEE), iniziato dal microbiologo Richard Lensky (Michigan State University) nel 1988. Egli ha osservato che un ceppo di E. coli, dopo decine di migliaia di generazioni, si è evoluto in modo da assumere la nuova e complessa capacità di metabolizzare il citrato. Il ceppo presentava anche divergenza riguardo la morfologia cellulare e la fitness, cioè il successo riproduttivo.
Anche la resistenza dei Batteri agli antibiotici, problema che si sta purtroppo facendo sempre più grave, è un esempio di rapido adattamento alle nuove condizioni ambientali, che dimostra come la selezione naturale sia sempre in azione.
Allo stesso modo si ha la resistenza degli Insetti ai pesticidi, la resistenza ai funghi patogeni, ai virus ecc.

Un esempio di selezione sessuale è fornito dagli esperimenti di John Endler su pesci della specie Poecilia reticulata, noti come guppy, molto usati negli acquari. La selezione spinge i maschi a modificarsi in due direzioni. Un gruppo che vive in ambienti con alto grado di predazione mantiene costantemente la colorazione arancione, perché i pesci con colorazioni rosse e gialle sono costantemente scartati dalle femmine e quindi hanno meno successo riproduttivo. Se, invece, la predazione è meno elevata, le femmine preferiscono i maschi grandi e dai colori appariscenti, anche se queste caratteristiche li rendono più attaccabili dai predatori.
