
Definizione
Secondo Darwin, gli individui che sopravvivono sono quelli "favoriti" da lievi variazioni vantaggiose. Questo processo di sopravvivenza dei "favoriti" fu chiamato da Darwin selezione naturale.
L'affermazione «sopravvivenza del più adatto» è stata tacciata di tautologia: «Chi è il più adatto? Quello che sopravvive. E chi sopravvive? Il più adatto». In realtà la selezione naturale favorisce alcuni organismi a danno di altri, senza alcun'altra specificazione. Nella maggior parte dei casi gli organismi favoriti sono più adatti all'ambiente nel quale vivono per certi aspetti, ma possono non esserlo per altri.
Un individuo può essere ben adattato al suo ambiente, ma se non si riproduce le sue caratteristiche "favorevoli" non vengono trasmesse alla discendenza perciò, in termini di genetica di popolazioni, la selezione naturale è ora definita come tasso differenziale di riproduzione di genotipi diversi in una popolazione, cioè la selezione naturale offre ai diversi individui di una data popolazione una diversa opportunità di lasciare una prole numerosa.
Una popolazione è sottoposta a selezione naturale quando uno o più genotipi hanno una maggiore o minore probabilità di sopravvivenza e/o di fecondità/fertilità rispetto agli altri genotipi. Se, infatti, un individuo possiede nel suo genotipo una combinazione di alleli favorevole, ha maggiori probabilità di sopravvivere e di riprodursi; di conseguenza, i suoi alleli hanno maggior probabilità di essere presenti in proporzione crescente nella generazione successiva. Se, invece, la combinazione degli alleli non è favorevole, l'individuo ha minori probabilità di sopravvivere e di riprodursi, e la presenza dei suoi alleli nella generazione successiva risulterà ridotta. In questo modo la popolazione diventa sempre più adatta, in termini di fertilità e sopravvivenza all'ambiente in cui vive.
La malattia, la competizione per le risorse, la predazione, i parametri ambientali sono i fattori che esercitano una selezione naturale sulla popolazione.
Il successo riproduttivo differenziale è il risultato delle interazioni tra i singoli organismi e il loro ambiente e l'accumulo nel tempo degli alleli favorevoli ha come risultato il cambiamento del pool genico e quindi l'evoluzione della popolazione.
Che cosa viene selezionato?
La selezione naturale agisce sull'individuo, o meglio, sull'intero fenotipo, che comprende tutti gli aspetti fisici, fisiologici e comportamentali riscontrabili in un organismo e non direttamente sul genotipo. Poiché il fenotipo è determinato geneticamente, la selezione può essere descritta in termini di variazioni di frequenze alleliche e genotipiche.
Un fenotipo generalmente è l'espressione di molti geni differenti, perciò ogni caratteristica fenotipica può essere ottenuta attraverso un certo numero di vie genotipiche differenti. Raramente un singolo allele può determinare un fenotipo vincente.
Il fenotipo, come abbiamo visto nelle pagine di genetica, è anche un prodotto dell'interazione del genotipo con l'ambiente. Per esempio, tra due gemelli umani identici si possono osservano, già al momento della nascita, alcune differenze, come il peso corporeo, dovute a differenze presenti all'interno dell'ambiente intrauterino.
Secondo Richard Dawkins, invece, l'unità fondamentale della selezione, non è la specie né il gruppo e neppure, in senso stretto, l'individuo, ma il gene.
Tipi di selezione naturale
Molti caratteri degli organismi sono poligenici, cioè controllati da molti geni e la variazione continua dei fenotipi è rappresentata con la curva "a campana", dove il fenotipo più comune si trova in posizione intermedia tra i due fenotipi estremi.
In base ai fenotipi possiamo avere tre tipi di selezione:
- Stabilizzante
- Direzionale
- Divergente
Selezione stabilizzante
La selezione stabilizzante, un processo sempre operante in tutte le popolazioni, comporta l'eliminazione degli individui con caratteri estremi favorendo quelli con valori medi; come risultato, la media della popolazione non cambia, ma la sua varietà diminuisce. Essa può aumentare l'adattamento della popolazione per quegli aspetti dell'ambiente che restano costanti.
Ad esempio, il peso medio dei bambini alla nascita è di circa 3 - 4 kg. I neonati che si discostano dalla media hanno una maggiore mortalità infantile: quelli troppo piccoli possono avere gli organi non del tutto funzionanti, mentre quelli con un peso maggiore possono portare a un parto tardivo e difficile.

Selezione direzionale
La selezione direzionale agisce in favore di individui con una caratteristica fenotipica estrema. Come risultato, cambia la media della popolazione, che si sposterà verso l'estremità che corrisponde al fenotipo più adatto. Essa, perciò, tende a sostituire gradualmente nel pool genico un allele (o un gruppo di alleli) con un altro. Tale situazione si riconosce quando una popolazione si sta adattando a un cambiamento dell'ambiente o quando migra in un ambiente diverso da quello di origine.
Un esempio di selezione direzionale è lo sviluppo della resistenza agli insetticidi.

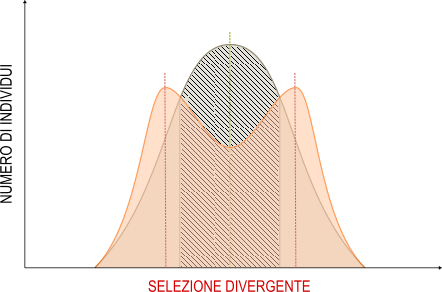
Selezione divergente
La selezione divergente (o disruptiva) induce un aumento della frequenza delle caratteristiche estreme di una popolazione a scapito delle forme intermedie. Come risultato cambiano le caratteristiche della popolazione aumentandone la variabilità. Si verifica quando una popolazione vive a cavallo tra due ambienti diversi.
Un esempio si ha per le piante che vivono al limite tra l'ambiente prativo e boschivo. Nelle aree più vicine al prato sopravvivono meglio quei fenotipi che sopportano gli ambienti aperti e luminosi, mentre nelle zone più vicine al bosco sono selezionati positivamente quei fenotipi che prediligono le zone ombrose. Con il passare delle generazioni, la popolazione potrà suddividersi in due sottopopolazioni distinte.
A questi tre tipi di selezione ne vanno aggiunti altri due:
- Selezione sessuale
- Selezione frequenza dipendente
La selezione sessuale
Alla selezione sessuale abbiamo già dedicato un paragrafo nella pagina di Darwin.
Si tratta di una selezione, conseguenza di un accoppiamento assortativo, in cui gli individui scelgono il partner in base a particolari caratteri secondari volti al successo riproduttivo, indipendentemente dalla capacità di sopravvivenza che tali caratteristiche potrebbero conferire.
Darwin trattò la selezione sessuale separatamente da quella naturale perché capì che si trattava di due meccanismi distinti, e talvolta contrastanti: mentre la selezione naturale favorisce i caratteri che aumentano la capacità di sopravvivenza, la selezione sessuale riguarda solo il successo riproduttivo.
Il principale agente della selezione sessuale, quindi, non è l'ambiente, ma il partner, che esercita il controllo diretto del successo riproduttivo scegliendosi il proprio compagno. I caratteri scelti, comunque, anche se possono apparire non adattativi, sembrano essere in relazione allo stato di salute del partner: per conservare caratteristiche così "costose", gli individui devono essere sani e vigorosi.

La selezione frequenza-dipendente
In questo tipo di selezione, diversamente dalle situazioni precedenti, il successo di un fenotipo dipende dalla frequenza con cui appare all'interno di una popolazione.
Nella selezione frequenza-dipendente negativa la selezione riduce la frequenza dei fenotipi più comuni e aumenta quelli meno comuni.
Nell'interazione preda-predatore si ha una selezione di questo tipo. Se, ad esempio un predatore può scegliere tra fenotipi diversi della stessa specie, si concentrerà su quello più comune, che progressivamente diminuirà la sua frequenza. Nello stesso tempo, il fenotipo raro ha la possibilità di prosperare e, a questo punto, il predatore cambierà obiettivo, mentre il primo fenotipo ha la possibilità di recuperare.
Anche nella competizione si ha una selezione frequenza-dipendente.
Questo tipo di selezione consente il mantenimento del polimorfismo all'interno della popolazione.
La selezione frequenza-dipendente positiva ha l'effetto opposto; favorendo forme comuni, tende ad eliminare variazione da una popolazione. Ad esempio, la presenza di forme particolarmente appariscenti all'interno della popolazione, anche se rare, potrebbe più facilmente attirare l'attenzione dei predatori e pertanto sarebbero selezionate negativamente, favorendo un ulteriore incremento dei fenotipi comuni.
La selezione agisce sul fenotipo, ma questo è l'espressione del genotipo pertanto, in relazione a quest'ultimo, possiamo distinguere quattro tipi di selezione.
- Selezione contro l'allele recessivo
- Selezione contro l'allele dominante
- Selezione in favore dell'eterozigote
- Selezione contro l'eterozigote
Selezione contro l'allele recessivo
Si presenta questa situazione quando gli individui di genotipo AA e Aa si riproducono con la stessa efficienza, mentre la capacità riproduttiva degli omozigoti aa è ridotta. L'effetto è la diminuzione della frequenza dell'allele recessivo, anche se sono necessarie molte generazioni per ridurlo sensibilmente.
Selezione contro l'allele dominante
Questo tipo di selezione è molto più efficace del precedente nell'eliminare l'allele sfavorito, in questo caso l'allele dominante.
Poiché, infatti, i genotipi AA e Aa hanno lo stesso fenotipo, la selezione agisce su entrambi. Al limite, se l'allele A è letale, basta una sola generazione per eliminarlo dal pool genico.
Selezione contro l'eterozigote
Si ha quando l'eterozigote è svantaggiato rispetto agli individui omozigoti.
Poiché ad ogni generazione alcuni individui eterozigoti non si riproducono, dal pool genico viene eliminato un uguale numero di alleli A e a e, di conseguenza, l'allele con frequenza minore è rimosso in percentuale proporzionalmente maggiore.
Se ho 100 alleli, di cui 10 sono a, l'eliminazione di un eterozigote Aa porta a una diminuzione di A dell'1,1% e di a del 10%.
Selezione in favore dell'eterozigote
In alcune situazioni gli individui eterozigoti possono avere un vantaggio nella sopravvivenza. In un paragrafo precedente l'abbiamo definito superiorità dell'eterozigote (o sovradominanza) e l'esempio proposto è quello dell'anemia falciforme.
La selezione a favore dell'eterozigote tende a mantenere entrambi gli alleli con frequenze abbastanza elevate nella popolazione (polimorfismo bilanciato).
Rispetto ai tre casi precedenti, è l'unico a mantenere la variabilità genetica, mentre gli altri tendono a ridurla.

Polimorfismo in
Dactylorhiza sambucina
La fitness
La fitness misura il successo riproduttivo di un organismo, cioè la capacità di trasmettere i propri geni alla generazione successiva in rapporto agli altri membri della popolazione in quel particolare ambiente e in quel particolare momento nel corso del tempo: un genotipo benefico in un determinato ambiente può essere dannoso in un altro. Il concetto di fitness non è sinonimo di adattamento ottimale all'ambiente.
La fitness è un parametro relativo, non assoluto: un organismo può avere maggiore o minore fitness relativamente agli altri membri della sua stessa popolazione.
La frequenza dei geni degli individui con fitness più elevata in un dato ambiente aumenterà da una generazione all'altra e quindi la frequenza degli alleli al locus interessato si modificherà, mentre la frequenza dei geni che determinano una bassa capacita riproduttiva diminuirà.
I componenti della fitness sono:
- sopravvivenza (mortalità differenziale),
- tasso di sviluppo,
- successo nell'accoppiamento,
- capacità di riprodursi (fecondità differenziale in termini di numero di gameti).
Per sopravvivere alla selezione, quindi, non basta essere più adatti, occorre essere anche più prolifici: un individuo forte e resistente, ma sterile, non contribuisce all'evoluzione della specie perché i suoi caratteri non vengono trasmessi. Inoltre, l'ambiente non fa sopravvivere gli individui, ma consente una differente prolificità (fitness).
La selezione in atto
Un classico esempio di selezione naturale a favore di un determinato genotipo in funzione di variazioni ambientali è quello definito melanismo industriale.
La Biston betularia è una farfalla notturna che trascorre il giorno riposandosi sui tronchi degli alberi o sulla superficie delle rocce.
Prima della rivoluzione industriale, in Inghilterra si osservavano numerosi esemplari di falena di colore chiaro, perfettamente adattati al colore della corteccia delle betulle e dei licheni grigi. Le falene scure erano rare.
Con l'avvento dell'industrializzazione, che usava grandi quantità di carbone, le superfici degli alberi e delle abitazioni si sono ricoperte di fuliggine che, insieme all'abbondante anidride solforosa, ha ucciso anche i licheni.
Le osservazioni fatte nel medesimo ambiente, dopo quarant'anni mostravano un consistente incremento delle forme melaniche, mentre quelle bianche erano quasi scomparse.
Mentre in precedenza le forme chiare potevano facilmente mimetizzarsi e sfuggire ai predatori, nel nuovo contesto ambientale sono le forme scure ad avere un vantaggio.
Negli ultimi decenni le emissioni di fuliggine si sono drasticamente ridotte, permettendo nuovamente la crescita dei licheni. Come previsto, le farfalle chiare sono in aumento e quelle scure diminuiscono.
Secondo Giuseppe Sermonti la favola delle farfalle è plausibile, ma non vera. Esistono, infatti, molti esempi di melanismo in luoghi senza industrie e le falene non hanno affatto l'abitudine di fermarsi sui tronchi, ma di nascondersi tra le fronde.

Biston betularia
(Crediti: E.B. Ford - [1964] 1975)
Il risultato della selezione naturale: l'adattamento
La selezione naturale è l'unico dei cinque agenti della microevoluzione che porta gli organismi all'adattamento all'ambiente.
Per adattamento si intendono quelle caratteristiche strutturali e comportamentali di una specie che aumentano la fitness, cioè le capacità di sopravvivenza e di riproduzione in un determinato ambiente.
L'adattamento non va confuso con la fitness perché esso è la conseguenza della selezione mentre la fitness è il diverso successo riproduttivo derivante dalla variabilità individuale. L'adattamento è il risultato di un processo di selezione che ha
perfezionato le primitive versioni dei tratti adattativi presenti nell'organismo attualmente vivente. L'individuo è da considerarsi ben adattato se riesce ad affermarsi nel corso del processo di selezione.
Il concetto di adattamento è relativo perché ciò che può essere adatto in un ambiente può non esserlo in un altro e, in seguito a variazioni ambientali, caratteristiche vantaggiose possono diventare svantaggiose e/o viceversa. Il processo di adattamento, perciò, garantisce la sopravvivenza a breve termine di una popolazione e, poiché l'ambiente si modifica in continuazione, sono richiesti continui aggiustamenti.

Nella Passera pianuzza
Platichthys flesus
gli occhi migrano durante lo sviluppo in modo da trovarsi sullo stesso lato del corpo
Un'idea sbagliata sulla selezione naturale è che alcune specie sono destinate all'estinzione perché "non si adattano" a un nuovo ambiente. Nelle popolazioni viventi l'evoluzione può verificarsi solo quando i tratti che influenzano la sopravvivenza sono già presenti in una popolazione e hanno una base genetica. Poiché la selezione agisce con il materiale che ha a disposizione e cioè le variazioni casuali presenti nel pool genico e l'espressione fenotipica che da questi ne deriva, il risultato non è mai perfetto, ma è un compromesso tra le esigenze adattative delle diverse caratteristiche di un organismo. Se si potesse partire ex novo con la creazione di un organo, il risultato sarebbe ottimale ma, dovendo modificare l'esistente, il risultato porta l'adattamento per quanto riguarda una caratteristica, ma spesso si presentano risvolti negativi per altri aspetti. È il caso della postura eretta dell'uomo, che comporta la tendenza ad avere il mal di schiena, il consumarsi dei dischi intervertebrali, la difficoltà nel parto, ecc.

Una straordinaria forma di adattamento è il mimetismo, come in questa falena
Adattamento all'ambiente fisico: clini ed ecotipi
Le popolazioni di una specie di organismi che si distribuiscono in un'ampia area geografica spesso sono divise in sottopopolazioni con flusso genico relativamente ridotto. Esse presentano differenze genetiche perché sottoposte a pressioni selettive legate alla graduale variazione delle caratteristiche ambientali, come temperatura, umidità, precipitazioni ecc.
La variazione graduale di uno o più caratteri ereditari lungo un asse geografico è detta cline.
Un cline presenta variazioni nell'espressione di caratteri, generalmente multifattoriali, come colore, dimensioni, forma, fisiologia o comportamento.
Ad esempio, le dimensioni del corpo di molti uccelli e mammiferi ampiamente distribuiti aumentano gradualmente all'aumentare della latitudine, presumibilmente perché gli animali più grandi sono in grado di resistere meglio alle temperature più fredde dell'inverno.
Le piante e gli animali che si distribuiscono lungo i clini presentano variazioni fenotipiche corrispondenti alle condizioni ambientali in cui vivono. Tali forme distinte sono dette ecotipi.

La
Platanthera bifolia
mostra normalmente un fenotipo slanciato con molti fiori lungo l'asse. Nelle aree alpine è invece più piccola, con fiori più raggruppati
Adattamento all'ambiente biologico: coevoluzione
La coevoluzione è l'evoluzione congiunta di organismi, legati fra loro da rapporti di reciproca dipendenza. Ciascuna popolazione interagisce così strettamente con l'altra tanto che si verificano adattamenti simultanei. Più in generale, tutti gli organismi evolvono simultaneamente agli altri con cui condividono l'ecosistema, perché nessuna specie vive totalmente isolata.
Queste interazioni comprendono i rapporti di tipo negativo come la competizione interspecifica o le relazioni che si instaurano tra predatore e preda o tra ospite e parassita, oppure rapporti mutualistici, in cui entrambe le specie interagenti beneficiano degli effetti della relazione.
Alcune caratteristiche dei fiori delle Angiosperme, ad esempio, si sono modificate in relazione agli insetti impollinatori; in alcuni casi, la forma del fiore e la posizione del nettario si adattano perfettamente all'apparato succhiatore dell'animale che, per nutrirsi, preleva involontariamente il polline da un fiore per deporlo poi su un altro della stessa specie. Anche le orchidee del genere Ophrys hanno modificato la forma e il colore del labello per farlo assomigliare all'insetto pronubo.

Impollinatore su
Anacamptis coriophora
-
Ophrys apifera
Il preadattamento
L'argomento del preadattamento è già stato trattato nella pagina di Darwin. Rivediamo qui i concetti secondo la biologia moderna.
Il problema riguarda il modo in cui la selezione riesce a portare alla formazione di strutture complesse che richiedono diversi stadi intermedi, quando questi non forniscono un vantaggio all'organismo che li possiede.
Possiamo individuare due modi per risolvere questo problema.
In alcuni casi le strutture complesse derivano da un processo di perfezionamento di strutture più primitive che svolgevano la medesima funzione.
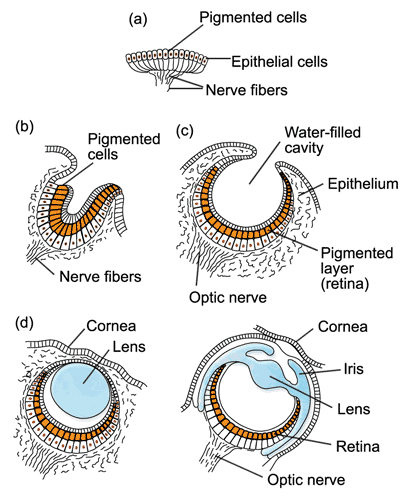
È il caso, ad esempio, dell'occhio.
Come aveva notato Darwin, gli occhi delle specie di Vertebrati sono estremamente complessi, essendo in grado di regolare la messa a fuoco, la quantità di luce e rilevare uno spettro di colori. Darwin ipotizzò che occhi così complessi dovessero evolversi da una struttura più semplice attraverso il processo della discendenza con modificazioni.
In origine abbiamo una semplice macchia oculare sulla pelle, costituita da due tipi di cellule, una cellula fotorecettrice e una cellula pigmentata adiacente. La cellula del fotorecettore, che è un tipo di cellula nervosa, è in grado di assorbire la luce e rispondere ad essa. La funzione della cella del pigmento è di impedire alla luce di raggiungere un lato della cella del fotorecettore. Questa disposizione primitiva a due celle consentirebbe a un organismo di percepire sia la luce che la direzione da cui proviene la luce. La selezione ha favorito quegli organismi in cui alcune cellule erano in grado di percepire la luce per rilevare il ciclo giorno-notte, l'ombra dei predatori o per localizzare il cibo. Occhi come questi si trovano ancora nelle specie moderne, come le larve di alcuni tipi di Molluschi.
I passaggi successivi portano un progressivo miglioramento delle strutture visive, differenziate in base ai gruppi di organismi. Le tappe comprendono lo sprofondamento dei recettori, che si trovano così riparati; la formazione di uno strato proteico protettivo trasparente e successivamente un ulteriore strato la cui variazione di spessore può diventare una specie di lente; successive modifiche consentono l'adattamento alla visione nei diversi tipi di ambiente.
Anche se gli occhi sono assai diversi nelle varie specie di organismi, sembra che ci sia un piano comune alla base di tutti i tipi di occhi, comprendente le opsine, molecole sensibili alla luce e alcuni geni, come i Pax-6, che guidano, durante lo sviluppo embrionale, la formazione di strutture in grado di catturare la luce, ma su questo argomento ci ritorneremo nell'ultima pagina.
La seconda via è quella che parte da strutture esistenti per adattarle a nuove funzioni, alternative a quella originaria, in una specie di bricolage.
In questo caso si parla di preadattamento o exattamento (exaptation): consiste nell'utilizzo, in mutate condizioni ambientali, di una struttura o di un comportamento per funzioni diverse rispetto a quelle originarie e solo in un secondo momento si modificano per adattarle alla nuova funzione.
Se gli organismi avessero ogni loro parte perfettamente progettata per svolgere un'unica funzione, l'adattamento in caso di variazioni ambientali non sarebbe possibile perché la modifica di una parte bloccherebbe il funzionamento dell'intero sistema. Si veda, come esempio, l'evoluzione degli uccelli in questa scheda.
Ulteriori dettagli ed esempi, come le penne degli uccelli, si trovano nella pagina di Darwin.
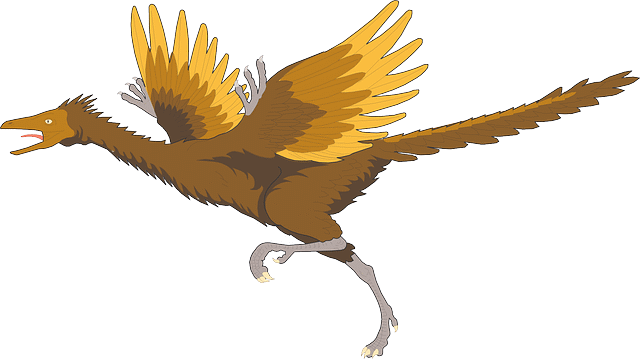
Archaeopteryx lithographica
La selezione produce organismi perfetti?
In natura non esiste l'organismo perfetto, ma solo l'organismo più adatto a sopravvivere in un ambiente definito e in un preciso periodo temporale.
L'evoluzione non è un progresso continuo verso una perfezione sempre maggiore, e questo per i seguenti motivi.
- Esistono dei limiti in quello che la selezione può fare, perché deve scegliere il più adatto tra gli organismi presenti in quel momento, e può non essere il più adatto in assoluto.
- I geni sono spesso pleiotropici perciò interessano diversi aspetti di un fenotipo e questo limita la possibilità di essere modificato perché potrebbero comparire contemporaneamente anche effetti che riducono la sopravvivenza.
- La selezione non crea nulla ma, in una specie di bricolage, rielabora le strutture già esistenti, ereditate dai suoi antenati, per adattarle a nuove situazioni.
- Gli adattamenti rappresentano spesso dei compromessi tra le diverse necessità dell'organismo Abbiamo già segnalato in precedenza che il bipedismo umano ha come conseguenza effetti negativi sulla colonna vertebrale.
- Poiché non c'è un finalismo lamarckiano per cui una struttura compare in base alle necessità, può non presentarsi mai la variazione genetica necessaria al perfezionamento di una determinata caratteristica.
- Le regole del gioco cambiano nel tempo. L'ambiente cambia in continuazione, perciò gli organismi più adattati in un certo momento, possono essere sfavoriti in un momento temporale diverso. Inoltre, la selezione potrebbe essere troppo lenta perché consenta agli individui di una popolazione ad adattarsi.
- Alcune soluzioni adottate possono condizionare, limitandola, l'evoluzione successiva. Per esempio, l'esoscheletro degli Artropodi ha consentito un'efficace difesa contro i predatori e la loro ampia diffusione. Questa struttura ha però limitato l'aumento delle dimensioni di questi organismi. I Vertebrati, invece, adottando un endoscheletro, risultano meno difesi, ma possono raggiungere dimensioni enormi.
- Il caso (deriva genetica) può consentire la sopravvivenza di gruppi di individui anche se non presentano caratteristiche più adatte rispetto ad altri della stessa specie e, in piccole popolazioni, può eliminare alleli che potrebbero essere migliori rispetto a quelli che si sono fissati.
- L'eccessiva specializzazione di alcune specie può limitare ulteriori sviluppi evolutivi. Le forme meno specializzate hanno maggiori possibilità di adattamento di fronte ai cambiamenti ambientali, mentre gli organismi troppo specializzati difficilmente hanno la possibilità di sopravvivere. Nel corso del tempo, le nuove forme che si susseguono derivano da antenati meno specializzati.
Con questi limiti non ci si può aspettare che l'evoluzione produca organismi perfetti. Osservando attentamente gli organismi viventi, si nota dunque un gran numero di "imperfezioni", spiegabili razionalmente solo con un processo evolutivo.
La teoria neutralista
In natura esistono geni con mutazioni che non modificano il funzionamento della proteina da essi codificata. All'inizio degli anni Sessanta si credeva che la selezione agisse sempre, anche sui questi geni mutati. In realtà, si è osservato che gli alleli neutrali, cioè quelli che non sono migliori o peggiori degli alleli alternativi per un dato locus, non modificano la fitness dell'individuo e sfuggono alla selezione naturale, accumulandosi nella popolazione.
Nel 1968 il biologo giapponese, Motoo Kimura (1924 - 1994), partendo da studi compiuti a livello molecolare, si accorse che il DNA muta con una velocità assai elevata, incrementando sensibilmente la variabilità genetica nelle sequenze amminoacidiche delle proteine e la maggior parte delle varianti sono selettivamente neutre. Questi polimorfismi quasi sempre non danno effetti fenotipici visibili e non si evidenzia alcuna correlazione con le condizioni ambientali.
Da queste considerazioni egli, insieme al suo collaboratore Tomoko Ohta, formulò la teoria neutralista o teoria neutrale dell'evoluzione, secondo la quale le mutazioni neutrali hanno un ruolo importante nei processi evolutivi. Esse si fissano nella popolazione in modo casuale piuttosto che per selezione darwiniana positiva, cioè la deriva genetica è più importante della selezione naturale, indipendentemente dalle dimensioni della popolazione perché, non alterando la fitness dell'organismo, le mutazioni neutrali non sono vagliate dalla selezione naturale.
Neutralità non significa che gli alleli siano tutti identici in termini di fitness. Significa invece che il destino di un allele è determinato in larga parte dalla deriva genetica casule. La selezione può agire, ma la sua intensità è troppo debole per sovrastare l'effetto del caso. La selezione naturale è ancora necessaria per spiegare l'adattamento; infatti, le rare mutazioni positive si affermano con la selezione e, allo stesso modo, quelle negative sono rapidamente da essa eliminate, ma rappresentano una minoranza di tutti i cambiamenti evolutivi che sono presenti e registrati nel DNA. Le mutazioni neutrali, invece, possono liberamente accumularsi e diffondersi in seguito alla deriva. Molti polimorfismi si mantengono perciò nella popolazione perché non sono controllati dalla selezione, in quanto essa agisce sul fenotipo e dipendono unicamente dalla loro comparsa per mutazione e dalla estinzione casuale.
Riassumendo, i principi cardine della teoria sono:
- la formazione di nuovi alleli mutanti neutrali;
- la fissazione degli alleli mutati nella popolazione tramite l'azione della deriva genetica.
Essi sono responsabili della maggior parte dei cambiamenti evolutivi e della variabilità all'interno delle specie.
Questa teoria suscitò molte polemiche da parte dei neodarwinisti ortodossi poiché sembrava inconciliabile con la visione darwiniana e con il ruolo della selezione naturale. In realtà sostiene soltanto che la selezione naturale agisce solo su quella bassa percentuale di mutazioni genetiche non neutrali in grado di produrre alterazioni, perciò oggi si ritiene che la teoria neutralista sia un contributo al neodarwinismo e un sostegno al ruolo della deriva genetica, assieme alla selezione naturale.