
Siamo arrivati alla fine di un lungo percorso evolutivo, iniziato 65 milioni di anni fa quando un gruppo di Insettivori si è avventurato sugli alberi alla ricerca di cibo. Da qui, nel tempo, si sono susseguiti vari adattamenti, come le estremità prensili, la verticalizzazione del corpo, ecc. I Primati si sono differenziati in diverse famiglie e generi che hanno colonizzato i vari continenti e da queste forme emergono gli Ominini. Per lungo tempo le Australopitecine hanno camminato accanto a noi, comparsi circa 2,8 milioni di anni fa, e poi, un milione di anni fa, siamo rimasti soli.
Il genere Homo, però, ha generato molti rami, spesso coesistenti, finché, 30.000 anni fa, H. sapiens rimane l'unica specie.
Questo genere si distingue dalle Australopitecine per le seguenti caratteristiche:
- bipedismo obbligato, ma non è stato il primo bipede;
- notevole aumento del volume cerebrale;
- predatore opportunista;
- ampia dispersione geografica;
- produttore di strumenti (inizia il Paleolitico), ma probabilmente non è stato l'unico.
Attualmente si conoscono le seguenti specie, tutte estinte tranne una.
- Homo habilis
- Homo rudolfensis
- Homo gautengensis
- Homo ergaster
- Homo georgicus
- Homo erectus
- Homo antecessor
- Homo heidelbergensis
- Homo neanderthalensis
- Homo denisovensis
- Homo floresiensis
- Homo luzonensis
- Homo naledi
- Homo sapiens
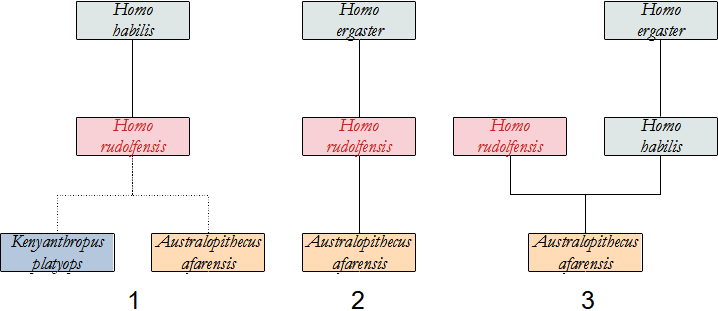
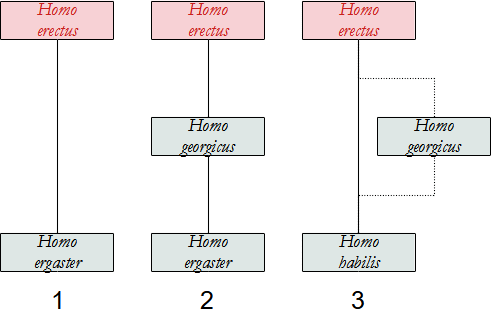
Assai discusso è il problema dell'individuazione di quale Ominino abbia dato origine al genere Homo, soprattutto dopo i recenti ritrovamenti.
L'ipotesi attualmente più accreditata vede in A. afarensis il progenitore di Homo, sia per motivi di ordine temporale, poiché è di poco antecedente a quest'ultimo, sia per motivi geografici.
Altri autori vedono in A. africanus il nostro antenato, ma ci sono problemi di tempo, perché il più antico reperto di Homo è antecedente ad A. africanus.
C'è anche chi propone Kenyanthropus platyops, di cui però abbiamo pochissime informazioni.
Qui sotto proponiamo uno schema delle possibili relazioni tra gli Ominini, che si presentano molto più intricate rispetto a quelle degli Australopitechi. Altri dettagli saranno forniti nella descrizione delle singole specie.

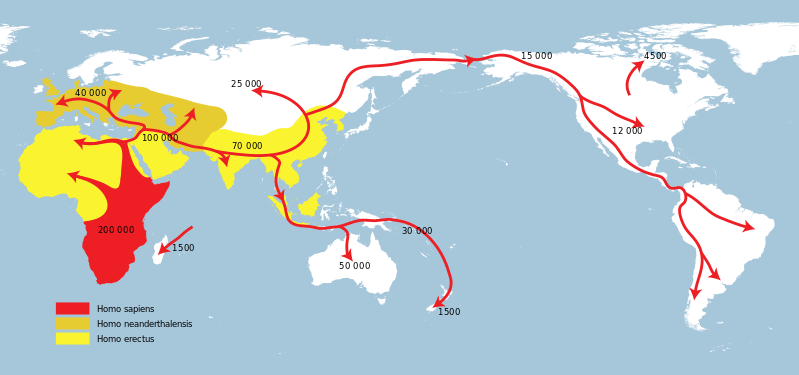
I rappresentanti della sottotribù Hominina non sono rimasti confinati nel continente africano perché a un certo punto della storia alcuni rappresentanti del genere Homo sono usciti dall'Africa e nell'arco di migliaia di generazioni hanno raggiunto l'Asia, l'Europa, l'Australia e il continente americano.
(Clicca sull'immagine per ingrandire)
Homo habilis
 Homo habilis, cioè "uomo capace, abile", perché ad esso sono stati attribuiti i primi strumenti in pietra dell'Olduvaiano, ma abbiamo già visto le problematiche all'attribuzione e alla datazione degli strumenti più antichi.
Homo habilis, cioè "uomo capace, abile", perché ad esso sono stati attribuiti i primi strumenti in pietra dell'Olduvaiano, ma abbiamo già visto le problematiche all'attribuzione e alla datazione degli strumenti più antichi.
Assai dibattuta è la sua collocazione, tra le Australopitecine o il genere Homo, perché presenta caratteristiche più avanzate rispetto Australopithecus, ma più primitive nei confronti di H. ergaster e anche per la grande variabilità dei resti, per cui è difficile definirne se si tratti di una o più specie.
I primi ritrovamenti risalgono agli inizi degli anni sessanta nella Gola di Olduvai in Tanzania. Successivamene sono stati individuati altri siti sia nell'Africa orientale (Etiopia, Kenya, Tanzania), sia nella Repubblica Sudafricana.
I reperti sono datati in un periodo compreso tra 2,4 a 1,44 milioni di anni fa, nel Pleistocene inferiore, ma una mandibola trovata nell'area di Ledi-Geraru nella regione di Afar è stata datata radiometricamente a 2,8 - 2,75 milioni di anni.
Ambiente e modo di vita
 H. habilis è vissuto in una zona piuttosto vasta dell'Africa orientale e del sud, in un ambiente ai margini della boscaglia, con praterie e foreste a galleria. Si pensa che frequentasse periodicamente luoghi ricchi di fonti d'acqua, di cibo e di materiale litico adatto a fabbricare strumenti.
H. habilis è vissuto in una zona piuttosto vasta dell'Africa orientale e del sud, in un ambiente ai margini della boscaglia, con praterie e foreste a galleria. Si pensa che frequentasse periodicamente luoghi ricchi di fonti d'acqua, di cibo e di materiale litico adatto a fabbricare strumenti.
H. habilis riusciva a convivere con le specie di Ominini contemporanei (H. rudolfensis, H. ergaster e P. boisei) probabilmente per la diversità di alimentazione. Pur avendo i denti ancora adatti a masticare alimenti duri, se ne cibava solo in caso di necessità, mentre in altri periodi prediligeva una varietà di vegetali teneri e frutti, insieme a carne ricavata da animali, forse non cacciati direttamente, dai quali strappava la carne con strumenti in pietra. H. habilis aveva, infatti, la capacità di fabbricare alcuni semplici utensili, costituiti da pietre e grossi ciottoli scheggiati a formare dei rozzi arnesi da taglio, comprendenti piccole schegge affilate, sfaldate da una pietra più grande, semplici pestelli e chopper, appartenenti all'industri litica dell'Olduvaiano (Paleolitico inferiore), che diedero a questo Ominino un vantaggio in un ambiente ostile, consentendogli l'accesso a risorse alimentari precluse ad altri animali.
H. habilis aveva, infatti, la capacità di fabbricare alcuni semplici utensili, costituiti da pietre e grossi ciottoli scheggiati a formare dei rozzi arnesi da taglio, comprendenti piccole schegge affilate, sfaldate da una pietra più grande, semplici pestelli e chopper, appartenenti all'industri litica dell'Olduvaiano (Paleolitico inferiore), che diedero a questo Ominino un vantaggio in un ambiente ostile, consentendogli l'accesso a risorse alimentari precluse ad altri animali.


Caratteristiche
La dubbia attribuzione di alcuni reperti fossili rende difficile avere un quadro preciso delle sue caratteristiche.
H. habilis è un Ominino abbastanza piccolo, alto mediamente 125 e 140 cm, con peso che varia tra 32 e 50 kg. L'elevato dimorfismo sessuale non è definitivamente confermato. Si ritiene che il corpo sia ancora ricoperto di peli.
 Il cranio è caratterizzato da una capacità di 500 - 750 cc, nettamente superiore, quindi, a quella degli Australopitechi; a ciò corrisponde a una volta cranica più alta e a una fronte più sviluppata. All'interno si notano le impronte delle due aree dell'emisfero cerebrale sinistro deputate all'espressione e alla comprensione del linguaggio e una maggiore ramificazione dei vasi sanguigni.
Il cranio è caratterizzato da una capacità di 500 - 750 cc, nettamente superiore, quindi, a quella degli Australopitechi; a ciò corrisponde a una volta cranica più alta e a una fronte più sviluppata. All'interno si notano le impronte delle due aree dell'emisfero cerebrale sinistro deputate all'espressione e alla comprensione del linguaggio e una maggiore ramificazione dei vasi sanguigni.
La testa è più tondeggiante rispetto a quella di Australopithecus/Paranthropus ed è priva della cresta sagittale per l'inserzione de muscoli masticatori; il foramen magnum è spostato in avanti.
Di contro, alcuni aspetti della faccia sono simili alle Australopitecine: è piatta ma ancora primitiva, con lieve prognatismo, un toro sopraorbitario presente ma non eccessivamente accentuato.
La mandibola è piuttosto leggera, ma non come quella degli umani moderni, e priva di mento osseo; l'arcata dentale è parabolica con denti posteriori ancora grandi rispetto all'uomo moderno, ma simili nella forma e nello spessore dello smalto. Anche gli incisivi sono grandi.
Le braccia sono ancora lunghe rispetto agli arti inferiori, in proporzione analoga a quella dello scimpanzé, perciò doveva trascorrere ancora parecchio tempo sugli alberi. A terra assumeva una locomozione bipede obbligata simile a quella dell'uomo moderno, con una certa capacità di corsa e di resistenza, in considerazione dell'alluce non più divergente e delle dita corte.
Le mani mostrano sia adattamenti all'arrampicata (falangi curve), sia la presa di precisione utile per la fabbricazione e manipolazione di strumenti.
Implicazioni evolutive
L'antenato di H. habilis, in base all'età dei reperti più antichi e ad alcuni aspetti morfologici, dovrebbe essere A. afarensis, direttamente o attraverso A. garhi, ma in questo caso sorgerebbero problemi di età e ancora di più con A. sediba. Proprio per l'età dovrebbe essere escluso anche H. rudolfensis. Ha attualmente minore credito anche l'origine da A. africanus.
Per quanto riguarda i discendenti, il candidato più accredito è H. ergaster. Sono stati tuttavia trovati reperti molto giovani di H. habilis e antichi di H. ergaster, perciò quest'ultimo non può essere un diretto discendente del primo. Si pensa allora, che ci sia un antenato comune ad entrambi oppure, per cladogenesi, da una popolazione di H. habilis si è formato il ramo di H. ergaster e i due hanno continuato a vivere parallelamente per almeno mezzo milione di anni.

Homo rudolfensis
 Homo rudolfensis è uno dei più antichi rappresentanti del genere Homo. L'esemplare tipo è stato scoperto dalla squadra di Richard Leakey nel 1972, a est del lago Turkana nel Kenia del nord. Il nome specifico deriva dalla vecchia denominazione del lago.
Homo rudolfensis è uno dei più antichi rappresentanti del genere Homo. L'esemplare tipo è stato scoperto dalla squadra di Richard Leakey nel 1972, a est del lago Turkana nel Kenia del nord. Il nome specifico deriva dalla vecchia denominazione del lago.
Il reperto è datato a 1,9 milioni di anni (Pleistocene inferiore), ma con i resti trovati nel 2007/09 abbiamo una datazione compresa tra 2,1 e 1,78 milioni di anni fa.
Se si aggiungono anche i resti trovati in Malawi, Tanzania, Kenya ed Etiopia, non attribuiti con certezza a H. rudolfensis, l'arco temporale si estende da 2,45 a 1,60 milioni di anni fa, periodo che coincide con la presenza nella stessa area di H. habilis, H. ergaster e P. boisei, mentre in Sudafrica c'è A. sediba.
L'inserimento in questo genere è dibattuto sia per le caratteristiche del cranio, sia per il numero limitato di reperti che non risolvono il problema della classificazione: una mascella e una mandibola con denti e un altro frammento di mandibola. Nessuna parte del corpo è stata trovata a parte due femori, che però non è provato che appartengano alla stessa specie del cranio.
Secondo alcuni autori le particolari caratteristiche del cranio grande e il volto lungo e piatto giustificano l'attribuzione della nuova specie Homo rudolfensis.
Per altri rientra nella variabilità di Homo habilis o forse si tratta di dimorfismo sessuale di questa specie e quindi H. habilis e H. rudolfensis sarebbero sinonimi.
Altri ancora lo fanno rientrare nella variabilità delle Australopitecine, con cui condivide alcuni caratteri, soprattutto dei denti, per cui dovrebbe essere Australopithecus rudolfensis.
Infine c'è chi nota affinità con Kenyanthropus platyops, perciò avremmo Kenyanthropus rudolfensis.
Ambiente e modo di vita
H. rudolfensis è endemico dell'Africa orientale e viveva nello stesso ambiente degli Ominini a lui contemporanei dove, nonostante i periodi di siccità, persistevano le foreste tropicali e i boschi.
La forma dei denti, se paragonata a quelli di altre specie, suggerisce che questo Ominino si cibasse di vegetali, forse più duri rispetto a H. habilis per la robustezza dell'apparato masticatorio e che mangiasse anche carne, in considerazione dell'aumento del volume cerebrale, che richiede un'ingente quantità di calorie.
H. rudolfensis potrebbe aver utilizzato strumenti di pietra per elaborare il cibo, ma non ne sono stati travati associati ad esso. Tuttavia sono stati trovanti strumenti litici dello stesso periodo in cui visse H. rudolfensis.
Caratteristiche
Il primo cranio ritrovato è abbastanza completo, ma senza alcuni denti, con un volume cerebrale valutato in circa 750 cc, grande più di quello di H. habilis, ma rapportato alla massa corporea non si discosta da quella dell'Australopiteco.

H. rudolfensis è più grande rispetto a H. habilis, ma secondo alcuni autori potrebbe trattarsi di maschi dello stesso H. habilis, ammesso che avessero un evidente dimorfismo sessuale. Si stima che i maschi fossero alti mediamente 160 cm e pesassero 60 kg, mentre le femmine 150 cm e 50 kg.

Differenza tra il cranio di
H. rudolfensis
, a sinistra, e
H. habilis
, a destra (Crediti:
Gunnar Creutz
- CC BY-SA 4.0)
_and_Homo_habilis_(KNM-ER_1813_cast)_at_G%C3%B6teborgs_Naturhistoriska_Museum_8595.jpg){kind=link}
Altre differenze con H. habilis sono: un piccolo toro sopraorbitale, una faccia molto più lunga, con la parte superiore più stretta in mezzo e prognatismo ridotto. Il mascellare è quadrato piuttosto che arrotondato. Non presenta la cresta sagittale di attacco dei muscoli masticatori, tipica del contemporaneo P. boisei. I molari e i premolari sono più grandi e con smalto spesso rispetto a H. habilis, simili a quelli delle Australopitecine, come simili sono i grandi zigomi, mentre gli incisivi sono più grandi di queste.

(Crediti:
Mauricio Antón
-
https://natuurtijdschriften.nl/pub/523523
- CC BY 3.0)
{kind=link}
Implicazioni evolutive
Per comprendere i legami filogenetici bisognerebbe avere un maggior numero di reperti, insieme a datazioni più sicure, al fine di una corretta collocazione tassonomica.
Alcuni autori ritengono che H. rudolfensis sia il discendente di K. platyops per le caratteristiche del cranio; altri lo ritengono discendente di A. afarensis.
Altrettanto discussa è la sua progenie. Mentre alcuni studiosi lo considerano l'antenato di H. habilis - ma i recenti ritrovamenti di quest'ultimo lo rendono improbabile per problemi di età -, altri lo vedono come un antenato diretto di H. ergaster; per altri ancora rappresenterebbe un ramo secco e sarebbe H. habilis l'antenato di H. ergaster.
Homo gautengensis
Homo gautengensis è un altro rappresentante del genere Homo non accettato da gran parte della comunità scientifica. Il nome specifico deriva dal luogo di ritrovamento, la provincia sudafricana Gauteng ("luogo dell'oro").
In questa specie sono inseriti alcuni fossili ritrovati in Sudafrica, precedentemente attribuiti a H. habilis, H. ergaster, A. africanus, P. robustus.
L'età dei reperti si estende per un periodo lunghissimo, compreso tra 1,8 e 0,8 milioni di anni fa.
Era alto circa un metro e pesava mediamente 50 kg e si differenzia da H. habilis soprattutto per i grandi denti atti alla masticazione dei vegetali, perciò probabilmente consumava meno carne di quest'ultimo.
Le braccia erano lunghe, perciò passava molto tempo sugli alberi ma camminava in posizione eretta, con andatura imperfetta.
Associati a questo Ominino sono stati trovati strumenti litici abbastanza primitivi e tracce di un possibile uso del fuoco.
Homo ergaster
 Homo ergaster è un Ominino vissuto in Africa nel Pleistocene inferiore. Il nome specifico deriva da "ἐργαστήρ", cioè "lavoratore" e quindi "uomo che lavora" perché associato a strumenti dell'industria litica dell'Acheuleano.
Homo ergaster è un Ominino vissuto in Africa nel Pleistocene inferiore. Il nome specifico deriva da "ἐργαστήρ", cioè "lavoratore" e quindi "uomo che lavora" perché associato a strumenti dell'industria litica dell'Acheuleano.
La specie fa riferimento ai fossili rinvenuti in Africa, distinti da Homo erectus, che invece popolava l'Asia, ma non tutti gli scienziati sono d'accordo. Essi ritengano che tutti i fossili asiatici e africani siano dell'unica specie Homo erectus, a causa delle limitate differenze, che esamineremo più avanti.
H. ergaster viveva nell'Africa orientale (Tanzania, Etiopia, Kenya) e nel Sudafrica in un periodo compreso tra 2 e 1 milione di anni fa. I reperti si concentrano soprattutto lungo le rive del lago Turkana, dove è stato trovato nel 1984 il famoso "ragazzo di Turkana", un esemplare di un giovane di circa 10 - 12 anni (ma forse 7 - 9, se aveva un ritmo di sviluppo veloce), datato a 1,6 milioni di anni fa, alto 160 cm, che da adulto avrebbe raggiunto 185 cm, altezza decisamente superiore rispetto a tutti gli Ominini incontrati in precedenza.
H. ergaster condivideva l'ambiente con H. habilis, H. rudolfensis, P. robustus, A. sediba.

Turkana boy (Crediti:
Neanderthal Museum
-
https://www.neanderthal.de/de/urmenschen.html
- CC BY-SA 4.0)
_Fundort_Nariokotome,_Kenia,_Rekonstruktion_im_Neanderthal_Museum.jpg){kind=link}
Ambiente e modo di vita
H. ergaster, rispetto agli Ominini precedenti, viveva in un ambiente di savana più aperta dove c'erano tanti pericoli, ma anche tanta carne, indispensabile per sviluppare e mantenere attivo un cervello che era aumentato di dimensioni. Per la prima volta abbiamo un comportamento di cacciatore-raccoglitore in cui non è più soltanto uno spazzino della savana, ma pratica la caccia attiva usando le pietre lavorate per colpire le prede. La carne però non è l'alimento preponderante ma si ciba di una grande varietà di alimenti. I denti non erano grandi come quelli dei predecessori perciò doveva ricavare l'energia necessaria da cibi di buona qualità, come piccoli animali, tuberi, bulbi, semi, noci e cibi duri che dovevano essere preparati con l'uso di strumenti.
I primi rappresentanti della specie sono associati alla tecnica olduvaiana, ma da 1,6 milioni di anni fa compare improvvisamente l'industria litica dell'Acheuleano, più raffinata della precedente, costituita da una grande varietà di strumenti e utensili comprendenti i bifacciali, che con la loro forma perfetta sono delle opere d'arte, strumenti più piccoli ricavati da schegge e asce a mano di varie dimensioni con funzioni differenziate.
Forse H. ergaster è stato il primo a controllare il fuoco per la cottura del cibo, rendendolo molto più digeribile. Ci sono tracce di un milione di anni fa in Sudafrica e forse ancora più antiche in Kenya.
Lo sfruttamento delle risorse di zone geografiche molto grandi, la stabilità di lunga durata nelle procedure di lavorazione della pietra potrebbero implicare lo sviluppo di un linguaggio complesso vocale e gestuale, possedendo probabilmente le strutture anatomiche per la fonazione.
Le capacità di costruire arnesi di pietra e di maneggiare il fuoco rese l'uomo in grado di distinguersi nettamente dalle altre specie di Primati.
Caratteristiche
Gli autori che distinguono H. ergaster da H. erectus hanno individuato sette caratteristiche distintive: una maggiore larghezza del cranio tra le ossa parietali; l'osso occipitale è più lungo; le ossa nasali più ampie; una maggiore apertura nasale; la base cranica è più ridotta; maggiore sviluppo della sinfisi mandibolare; molari più stretti e canini più corti. Non tutti però concordano su queste analisi.
Altre caratteristiche saranno descritte con H. erectus.
H. ergaster presenta un aumento significativo sia nell'altezza che nella massa corporea, assumendo proporzioni più simili ai membri successivi del genere Homo e quindi distanti dagli Australopitechi: l'altezza è compresa tra 145 e 190 cm, con peso 52 - 65 kg e il dimorfismo sessuale è relativamente inferiore rispetto ai precedenti Ominini. Il volume endocranico è aumentato fino a 850 - 900 cc, ma ciò non implica una maggiore intelligenza perché la crescita del cervello è proporzionale a quella del corpo, perciò l'aumento non è particolarmente significativo rispetto a H. habilis.
Il volume endocranico è aumentato fino a 850 - 900 cc, ma ciò non implica una maggiore intelligenza perché la crescita del cervello è proporzionale a quella del corpo, perciò l'aumento non è particolarmente significativo rispetto a H. habilis.
Il cranio acquista un'arcata sovraorbitale abbastanza robusta, molto probabilmente il risultato dei lobi frontali ed occipitali espansi, mentre la volta cranica era lunga e bassa, perciò la fronte è ancora sfuggente. Anche la regione nucale è pesante.
Le ossa nasali suggeriscono un naso sporgente, con le narici orientate verso il basso, simile a quello umano, come adattamento alle zone aride.
Continua la riduzione della dentizione e dell'apparato masticatorio, che sono più piccoli di quelli di H. habilis e H. rudolfensis, indicando un cambiamento importante nella dieta. In particolare, pur avendo la mandibola robusta e le mascelle sporgenti in avanti, i molari sono abbastanza piccoli. Il mento è assente.
Il torace comincia ad assumere una forma più a botte, con fianchi stretti e le gambe sono relativamente lunghe, anatomicamente simili a quelle degli esseri umani moderni, che li avrebbero resi obbligatamente bipedi, mentre le braccia non sono più lunghe delle gambe, ma di più rispetto alle attuali. Questo lo rende diverso dagli Ominini precedenti, che combinano il bipedismo all'arrampicata sugli alberi.
L'adattamento all'ambiente arido e caldo avrebbe portato H. ergaster ad avere, il primo del genere Homo, la pelle quasi glabra, nuda e molto scura, almeno in Africa, perché se si è spostato nelle fredde regioni dell'Eurasia doveva avere una copertura di peli per sopravvivere, a meno che non usasse il fuoco per scaldarsi o si coprisse in qualche modo.

(Crediti:
Mauricio Antón
, published with Alan Turner -
Africa – the Evolution of a Continent and its Large Mammal Fauna
- CC BY 4.0)
{kind=link}
Implicazioni evolutive
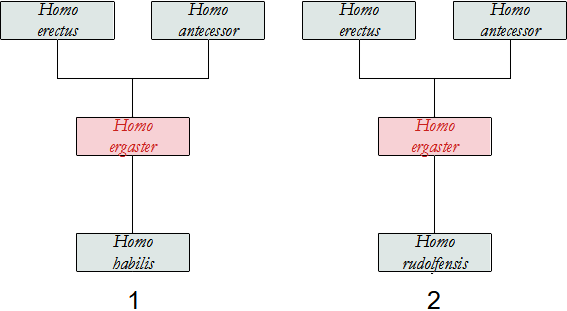
H. ergaster deriverebbe da H. habilis o da H rudolfensis per un processo di cladogenesi, perché c'è un ampio intervallo si tempo in cui le specie convivono.
Per quanto riguarda i discendenti, sarebbe l'antenato diretto di H. erectus in Asia, di H. heidelbergensis e di H. sapiens in Africa, di H. antecessor in Europa, di H. georgicus nella Repubblica di Georgia ed eventualmente H. floresiensis sull'isola di Flores.
Migrazioni
H. ergaster fu il primo (forse) rappresentate del genere Homo a uscire dall'Africa: Out of Africa I. I piedi lunghi e le aumentate dimensioni del cervello hanno fatto di questo Ominino un tipo adatto all'espansione: i fossili suggeriscono che si è diffuso dall'Africa nord-orientale all'Asia centrale ed orientale e forse anche in Europa del sud (Apricena [FG] - 1,5 m.a.), sviluppando nel tempo caratteristiche proprie che hanno originato le diverse specie.
Si suppone che gli spostamenti siano avvenuti molto precocemente, forse già 2 milioni di anni fa, portando con sé la primitiva industria litica olduvaiana. Ci sono tuttavia pietre lavorate provenienti dalla Cina vecchie di 2,1 milioni di anni e quindi o si retrodata la comparsa di H. ergaster o è stato H. habilis il primo a lasciare l'Africa.
H. ergaster lo troviamo molto presto in Medio Oriente a Ubediya e poco dopo, 1,77 - 1,85 milioni di anni fa, lo troviamo a Dmanisi in Georgia come H. georgicus, che sicuramente proveniva dall'Africa.
Forse già un milione 900 mila anni fa lo troviamo in Pakistan e poi arriva anche in Cina, dando origine a H. erectus.
L'espansione si divide in due rami, quello settentrionale, l'Uomo di Pechino e meridionale, l'Uomo di Giava.
Sempre dal Medio Oriente i primi umani arriveranno anche in Europa, dando origine a un'altra varietà geografica che è H. antecessor.
Non dobbiamo pensare a migrazioni in senso moderno, ma piuttosto a un'espansione progressiva, avvenuta nell'arco di migliaia di generazioni, man mano che si estendeva la savana, inseguendo gli animali di cui si nutrivano, quando le risorse locali si esaurivano o quando la popolazione aumentava tanto da non essere in grado di sostentarsi, soprattutto se c'era competizione con altri Ominini.
Il passaggio nel continente Eurasiatico può essere avvenuto attraversando il Mar Rosso o risalendo la valle del Nilo o anche attraversando il Mare Mediterraneo quando c'erano importanti abbassamenti del livello marino.

(Clicca sull'immagine per ingrandire)
Homo georgicus
Tra il 1991 e il 2001 sono stati trovati a Dmanisi, in Georgia, resti di un Ominino datati radiometricamente 1,8 milioni di anni fa (Pleistocene inferiore), che è stato chiamato Homo georgicus. È il più antico rappresentante del genere Homo rinvenuto in Eurasia, anche se ci sono strumenti più vecchi rinvenuti in Cina e si presume che, partendo dall'Africa, sia arrivato nel Caucaso meridionale attraverso la Palestina.
L'assegnazione a una nuova specie non è condivisa dalla comunità scientifica, che lo ritiene un rappresentante di H. ergaster/erectus.

(Crediti:
Cicero Moraes et alii
(Luca Bezzi, Nicola Carrara, Telmo Pievani) -
http://arc-team-open-research.blogspot.it/2015/10/homo-georgicus.html
- CC BY 4.0)
{kind=link}
Caratteristiche
Nel sito georgiano sono stati trovati fossili di 5 crani, 4 mandibole, parecchi denti isolati e molti resti postcraniali di individui appartenenti a diverse fasce d'età.
Le caratteristiche sono intermedie tra quelle di H. habilis e H. ergaster: cranio robusto, capacità cranica tra 500 e 750 cc, faccia prognatica, mancanza del naso sporgente, molari grandi e canini lunghi.

Gli arti superiori sono simili a quelle dell'Australopiteco, segno che si arrampicava ancora agevolmente sugli alberi per rifugiarsi o per cercare cibo. Le gambe invece sono più moderne e in grado di compiere lunghi percorsi.
La taglia è piccola, 145 - 165 cm, con elevato dimorfismo sessuale.
Ambiente e modo di vita
 H. georgicus viveva in ambiente di savana più umida di quella africana e ricca d'acqua, con macchie boscose e un clima simile a quello mediterraneo. In questo ambiente trovava i vegetali di cui si nutriva, spesso fibrosi e di bassa qualità.
H. georgicus viveva in ambiente di savana più umida di quella africana e ricca d'acqua, con macchie boscose e un clima simile a quello mediterraneo. In questo ambiente trovava i vegetali di cui si nutriva, spesso fibrosi e di bassa qualità.
Associati a questo Ominino sono stai trova strumenti dell'industria litica dell'Olduvaiano.
Implicazioni evolutive
Alcuni studiosi ritengono che H. georgicus sia un discendente di H. ergaster o una variazione locale dello stesso, migrata precocemente nel Caucaso, da cui deriverebbe H. erectus asiatico.
Altri ritengono che sia più vicino a H. habilis, uscito prima di H. ergaster dall'Africa, evoluto in H. georgicus e trasferito in Africa diventando H. ergaster e in Asia evolvendo in H. erectus.
Noi riteniamo che una porzione di popolazione assai primitiva di H. ergaster, (pre-ergaster) con ancora caratteristiche di H. habilis, sia migrata in Georgia dando origine a H. georgicus, mentre la parte di popolazione rimasta in Africa ha originato H. ergaster. Questa proposta è simile alla precedente ma giustifica l'origine africana.

Homo erectus
 Homo erectus è un Ominino scoperto nel 1891 nell'isola di Giava. Inizialmente denominato "Uomo di Giava" o Pithecanthropus erectus, cioè "uomo-scimmia eretto", perché lo si riteneva l'anello di collegamento tra la scimmia e l'uomo, i cui antenati erano ritenuti ricurvi e poi lentamente diventati eretti, è stato ascritto poi al genere Homo insieme all'Uomo di Pechino, Sinanthropus pekinensis (sinantropo = insieme all'uomo) trovato in Cina. I reperti cinesi appaiono più moderni di quelli di Giava, segno di una seconda ondata migratoria più recente.
Homo erectus è un Ominino scoperto nel 1891 nell'isola di Giava. Inizialmente denominato "Uomo di Giava" o Pithecanthropus erectus, cioè "uomo-scimmia eretto", perché lo si riteneva l'anello di collegamento tra la scimmia e l'uomo, i cui antenati erano ritenuti ricurvi e poi lentamente diventati eretti, è stato ascritto poi al genere Homo insieme all'Uomo di Pechino, Sinanthropus pekinensis (sinantropo = insieme all'uomo) trovato in Cina. I reperti cinesi appaiono più moderni di quelli di Giava, segno di una seconda ondata migratoria più recente.





Le non facili datazioni radiometrica indicano un'età compresa tra 1,8 milioni di anni (Pleistocene) e 117/108.000 anni fa, quindi una specie assai longeva: quasi 2 milioni di anni!
In questo paragrafo è considerato come specie asiatica, distinta dall'H. ergaster africano.
H. erectus è stato trovato anche in Africa (Tanzania, Etiopia), in depositi di 1,5 - 1,2 milioni di anni fa e si pensa che sia una migrazione dall'Asia.
Ambiente e modo di vita
 H. erectus si è espanso su ampi territori del continente asiatico che presentavano climi differenziati. Le sue capacità intellettive gli hanno consentito di essere meno dipendente dall'ambiente, creando le prime capanne nei luoghi sprovvisti di rifugi naturali o sfruttando le caverne che illuminava grazie all'uso del fuoco, che serviva anche per cuocere i cibi, per riscaldarsi e per difendersi dagli animali che penetravano nelle caverne. Molti studiosi ritengono tuttavia che l'uso del fuoco fosse raro, sporadico e opportunistico da parte di questa specie.
H. erectus si è espanso su ampi territori del continente asiatico che presentavano climi differenziati. Le sue capacità intellettive gli hanno consentito di essere meno dipendente dall'ambiente, creando le prime capanne nei luoghi sprovvisti di rifugi naturali o sfruttando le caverne che illuminava grazie all'uso del fuoco, che serviva anche per cuocere i cibi, per riscaldarsi e per difendersi dagli animali che penetravano nelle caverne. Molti studiosi ritengono tuttavia che l'uso del fuoco fosse raro, sporadico e opportunistico da parte di questa specie.
Era un buon cacciatore di antilopi e di altri animali, dei quali, oltre al cibo, aveva imparato a sfruttare le pelli per difendersi dal freddo. La sua dieta però non era solo di natura proteica, ma si nutriva anche di una grande varietà di vegetali.
La necessità di procurarsi cibo e di costituire delle riserve per i periodi poco produttivi, fu forse il primo impulso alla cooperazione all'interno del gruppo e alla divisione dei compiti, che permisero certamente di cacciare, con più probabilità di successo, animali anche di dimensioni maggiori. La cooperazione si estende anche nella cura dei membri del gruppo feriti o malati, che non erano in grado di procacciarsi il cibo in modo autonomo.
A H. erectus è associata l'industria litica dell'Acheuleano, che giunge 1 milione di anni fa dopo una seconda espansione dall'Africa, costituita da strumenti e utensili più perfezionati rispetto alla tecnica precedente, che è stata comunque utilizzata nel primo periodo, e dall'impiego della selce. Con questa roccia vetrosa era in grado di costruire arnesi più affilati e taglienti, che consentivano di scuoiare e tagliare a pezzi gli animali uccisi, rendendo più facile il trasporto e la divisione del cibo fra i componenti del gruppo. L'esecuzione molto accurata e le evidenti simmetrie di quegli utensili, come le amigdale, denunciano nei loro costruttori non soltanto l'attenzione alla funzionalità dell'oggetto, ma anche una sensibilità estetica.
La migrazione in aree lontane, la cooperazione nella caccia e la costruzione di utensili così perfezionati richiede un'efficace modalità di comunicazione. Non sappiamo se avesse tutte le strutture anatomiche per parlare, anche se erano presenti le aree cerebrali deputate al linguaggio, però si ipotizza che usasse una specie di protolinguaggio, non paragonabile a quello dell'uomo moderno.
Caratteristiche
 Il corpo di H. erectus si avvicina notevolmente alle nostre dimensioni, peso 45 - 70 kg, altezza 145 - 185 cm, con un dimorfismo sessuale ancora evidente ma che si sta avvicinando a quello degli uomini moderni.
Il corpo di H. erectus si avvicina notevolmente alle nostre dimensioni, peso 45 - 70 kg, altezza 145 - 185 cm, con un dimorfismo sessuale ancora evidente ma che si sta avvicinando a quello degli uomini moderni.
Il cranio si differenzia dagli altri Ominini, non tanto per la capacità cranica, che è di 820 cc nei più antichi, mentre quelli più evoluti arrivano mediamente a 1050 cc, non molto superiore a quella di H. ergaster, ma nella forma lunga e bassa. La fronte si alza ancora e la calotta si espande: nell'H. erectus si espande all'indietro, nell'H. sapiens verso l'alto.
La faccia è piatta e il naso sporgente come l'uomo attuale, si accompagnano ad aspetti profondamente diversi, come il grande rilievo del toro sopraorbitale, un toro nucale, un leggero prognatismo e un mento in pratica inesistente.

(Crediti:
James St. John
- CC BY 2.0)
_%26_indochinite_tektites_(Pleistocene;_southeastern_Asia)_3_(45586418921).jpg){kind=link}
Si osserva una riduzione della dentatura posteriore (ma più grande di quella odierna), correlata a una diminuzione nella mandibola e lo smalto dei denti è sottile.
Lo scheletro è quasi uguale, ma più robusto, a indicare una grande resistenza; la cassa toracica ha una forma più cilindrica; si nota una riduzione dell'avambraccio e le gambe dimostrano che la sua andatura è molto simile alla nostra, molto efficiente nelle lunghe distanze. Questo è dimostrato dalle orme di almeno 20 individui lasciate in Kenya 1,5 milioni di anni fa e quelle dell'Eritrea di 800.000 anni fa.
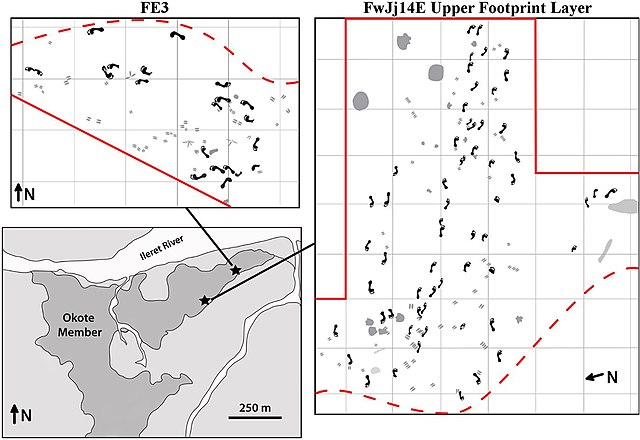
Impronte trovate a Ileret in Kenya (Crediti:
Hatala, K. and colleagues
-
https://doi.org/10.1038/srep28766
- CC BY-SA 4.0)
{kind=link}
Implicazioni evolutive
H. erectus secondo alcuni scienziati è il discendente diretto di H. ergaster da cui si sarebbe separato 1,6 milioni di anni fa, o di H. georgicus.
Per altri H. habilis, arrivato molto precocemente in Asia, si è evoluto localmente in H. erectus e poi diffuso nella parte orientale (Uomo di Pechino e Uomo di Giava) fino a giungere in Africa e forse in Europa. Le relazioni filogenetiche con H. sapiens e gli altri Ominini europei le esamineremo più avanti.
Homo antecessor
Homo antecessor, che in latino significa "esploratore", "pioniere" o "predecessore" è una specie indicata sulla base di 80 reperti di 6 individui, ritrovati nel 1994-95 nei due siti Gran Dolina ad Atapuerca, nel nord della Spagna, datati radiometricamente a 780.000 anni fa, rendendoli tra i più vecchi Ominini europei conosciuti. I ritrovamenti a Sima del Elefante ("Fossa dell'Elefante") del 2007 estendono la datazione a 1,2 milioni di anni fa (Pleistocene inferiore).
Gli Ominini sono esemplari giovani: "il figlio della Gran Dolina" sembra avere 10 - 11 anni, mentre gli altri hanno un'età compresa tra i 20 e 25 anni.

(Crediti: Clicca sull'immagine - CC BY-NC-ND 4.0)
Ambiente e modo di vita
H. antecessor viveva nelle zone boscose della penisola iberica, in un periodo in cui il clima, nelle sue continue oscillazioni, doveva essere caldo e umido. Quando il clima divenne più freddo, venne sostituito da nuove specie meglio adattate alle nuove condizioni.
Associato ai suoi fossili sono stati trovate ossa di giovani orsi, cavalli, bisonti, iene, linci, cinghiali, roditori, varie specie di cervo, di cui probabilmente si cibava, insieme ad alimenti di natura vegetale. Molto importante nella dieta è il midollo osseo ottenuto grazie all'impiego di utensili. Accanto ai reperti sono stati, infatti, trovati circa 250 strumenti appartenenti all'industria litica dell'Acheuleano, le cui tracce d'uso sono state osservate in quasi meta delle ossa umane, particolarmente del cranio, segno che si praticava il cannibalismo, soprattutto in individui giovani. Probabilmente la carne umana entrava nella dieta quando scarseggia altro tipo di cibo e non sembra riferirsi a qualche rituale funerario. Alcuni autori ritengono invece che l'area non scarseggiasse di cibo e il cannibalismo era praticato in modo sistematico nei confronti delle tribù vicine e tribali.
Non sembra che conoscesse l'uso del fuoco.
Caratteristiche
 H. antecessor mostra una miscela di caratteristiche arcaiche di H. ergaster e moderne di H. heidelbergensis e H. sapiens.
H. antecessor mostra una miscela di caratteristiche arcaiche di H. ergaster e moderne di H. heidelbergensis e H. sapiens.
Il corpo ha una struttura simile all'uomo moderno, ma più robusto, con un'altezza di 160 - 180 cm e il peso che poteva arrivare a 90 kg.
Il cranio ha una capacità di circa 1.000 cc, ma potrebbe essere di una femmina o di un individuo giovane. La faccia è appiattita e il naso sporgente, ma la fronte è bassa e marcata, con un rilievo nucale; i denti assomigliano a quelli di H. ergaster, con grandi canini e incisivi.

Implicazioni evolutive
I pochi resti presenti hanno sempre reso difficoltoso formulare ipotesi serie sull'origine di H. antecessor e forse sarebbe opportuno attendere nuove scoperte.
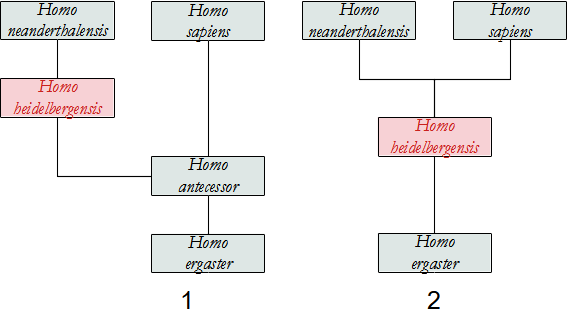
Gli scopritori di questo Ominino ritengono che da una popolazione di H. ergaster si sarebbe separato un gruppo che si è espanso verso l'Asia, originando H. erectus (si veda questo Ominino per i dettagli). Un gruppo, sempre di H. ergaster, già in Africa si sarebbe evoluto in H. antecessor e poi sarebbe migrato in Europa circa 800.000 anni fa, passando attraverso il Medio Oriente, anche se al momento non abbiamo prove fossili della sua esistenza in Africa. In quest'area si sarebbe evoluto H. heidelbergensis e da questo H. neanderthalensis. La parte di popolazione di H. antecessor rimasta in Africa avrebbe dato origine a H. sapiens che in seguito ha soppiantato tutte le altre specie. Secondo questa ipotesi, dunque, H. antecessor è l'ultimo antenato comune tra H. neanderthalensis e sapiens.
Un'altra ipotesi vede invece H. heidelbergensis l'antenato comune tra neanderthalensis e sapiens, mentre H. antecessor, se è specie valida e non una variazione locale di H. heidelbergensis o di H. ergaster/erectus, avrebbe avuto origine direttamente in Europa Meridionale.
Nel 2020 un gruppo di Copenaghen è riuscito a studiare il proteoma ricavato da un molare. Dall'analisi emerge che H. antecessor è uno stretto parente dell'ultimo antenato comune a H. sapiens, neanderthalensis e Denisova, non l'ultimo antenato comune ma un "gruppo gemello" posto alla base di questi.

Homo heidelbergensis
 Homo heidelbergensis, precedentemente chiamato Homo sapiens arcaico, riguarda un gruppo di vari crani che presentano caratteristiche di H. erectus, di H. neanderthalensis e di esseri umani moderni. Precisiamo che in questo Ominino vengono racchiusi reperti denominati Homo rhodesiensis, trovato in diversi siti Africani; alcuni autori includono anche H. antecessor, che invece noi abbiamo trattato separatamente. Ciò deriva dal fatto che la comunità scientifica non si è messa ancora d'accordo, né c'è una visione maggioritaria del problema.
Homo heidelbergensis, precedentemente chiamato Homo sapiens arcaico, riguarda un gruppo di vari crani che presentano caratteristiche di H. erectus, di H. neanderthalensis e di esseri umani moderni. Precisiamo che in questo Ominino vengono racchiusi reperti denominati Homo rhodesiensis, trovato in diversi siti Africani; alcuni autori includono anche H. antecessor, che invece noi abbiamo trattato separatamente. Ciò deriva dal fatto che la comunità scientifica non si è messa ancora d'accordo, né c'è una visione maggioritaria del problema.

 L'esemplare tipo è stato scoperto nel 1907 nelle cave di sabbia di Mauer vicino ad Heidelberg, in Germania e da qui il nome specifico. L'età esatta dell'esemplare è incerta, ma dovrebbe aggirarsi intorno ai 500.000 anni. Anche a Bodo d'Ar, in Etiopia è stato trovato un cranio che potrebbe essere attribuito a H. heidelbergensis, così come ci sono resti nella caverna di Arago, a Tautavel, in Francia, nelle montagne orientali dei Pirenei.
L'esemplare tipo è stato scoperto nel 1907 nelle cave di sabbia di Mauer vicino ad Heidelberg, in Germania e da qui il nome specifico. L'età esatta dell'esemplare è incerta, ma dovrebbe aggirarsi intorno ai 500.000 anni. Anche a Bodo d'Ar, in Etiopia è stato trovato un cranio che potrebbe essere attribuito a H. heidelbergensis, così come ci sono resti nella caverna di Arago, a Tautavel, in Francia, nelle montagne orientali dei Pirenei.
In Italia, in provincia di Caserta, ci sono le Ciampate del Diavolo, impronte fossili su cenere lavica di circa 350.000 anni fa.

Ciampate del Diavolo (Crediti:
edmondo gnerre
- CC BY 2.0)
{kind=link}
 Sempre in Italia, nel sito di Isernia La Pineta (Molise), è stato scoperto nel 1979 un paleosuolo coperto di ossa - alcune spaccate - di Bisonte, Elefante antico, Rinoceronte, Orso, ecc., insieme a numerosi utensili (chopper, raschiatoi, denticolati in selce), datato radiometricamente a 736.000 anni fa. Nel 2014 è stato trovato un incisivo superiore di bambino appartenente a una variazione locale di H. heidelbergensis, chiamato Homo aeserniensis, con un'età stimata di 586.000 anni.
Sempre in Italia, nel sito di Isernia La Pineta (Molise), è stato scoperto nel 1979 un paleosuolo coperto di ossa - alcune spaccate - di Bisonte, Elefante antico, Rinoceronte, Orso, ecc., insieme a numerosi utensili (chopper, raschiatoi, denticolati in selce), datato radiometricamente a 736.000 anni fa. Nel 2014 è stato trovato un incisivo superiore di bambino appartenente a una variazione locale di H. heidelbergensis, chiamato Homo aeserniensis, con un'età stimata di 586.000 anni.
I reperti indiani e cinesi sono controversi perché alcuni studiosi ritengono debbano essere considerati solo fossili europei e discussi sono anche quelli della Gran Dolina, in Spagna, dove, come abbiamo visto, sono in continuità con i resti di H. antecessor, vissuto 200.000 anni prima.
Esemplari sono stati trovati anche in Ungheria, Grecia, Marocco, Zambia, Tanzania, Sud Africa, Cina.
H. heidelbergensis è vissuto in un periodo compreso tra 600.000 e 100.000 anni fa, nel Pleistocene medio, ma potrebbe essere già comparso tra un milione e 800.000 anni fa in Africa, secondo le analisi del DNA ed è il protagonista della migrazione Out of Africa II.

(Clicca sull'immagine per ingrandire)
Ambiente e modo di vita
 H. heidelbergensis viveva in un periodo di oscillazioni climatiche e durante una fase interglaciale si è trasferito dall'Africa in Europa, dove il clima era più freddo. Il precedente H. antecessor, meno adattato alle zone fredde, è stato sostituito da un Ominino più robusto e in grado di difendersi meglio dalle basse temperature, probabilmente coprendosi con gli animali che cacciava.
H. heidelbergensis viveva in un periodo di oscillazioni climatiche e durante una fase interglaciale si è trasferito dall'Africa in Europa, dove il clima era più freddo. Il precedente H. antecessor, meno adattato alle zone fredde, è stato sostituito da un Ominino più robusto e in grado di difendersi meglio dalle basse temperature, probabilmente coprendosi con gli animali che cacciava.
Sapeva usare il fuoco ma non ci sono prove sicure di un uso costante; si riparava in grotte naturali e forse costruiva semplici rifugi in legno e pietra, come quello rinvenuto a Terra Amata in Francia.
Era un abile cacciatore di animali, forse anche di grossa taglia, come provano le ossa di rinoceronte, orso, elefante, ippopotamo, bisonti, cervi e soprattutto cavalli, che uccideva con lance in legno oppure con i bifacciali litici dell'industria dell'Acheuleano.
Le strategie di caccia che richiedevano cooperazione, il riunirsi attorno al fuoco, la trasmissione delle tecniche di lavorazione degli strumenti litici, nonché le strutture anatomiche, provano che H. heidelbergensis usava una qualche forma di linguaggio primitivo.
(Crediti: Clicca sull'immagine - CC BY-NC-ND 4.0)
 Si ritiene anche che seppellisse i morti con una specie di rito religioso o sacrificale, sulla base della trentina di scheletri trovati nella "Fossa delle ossa" ad Atapuerca, insieme a un'ascia in quarzo rosa, ma potrebbe trattarsi di un seppellimento per impedire l'attacco di animali.
Si ritiene anche che seppellisse i morti con una specie di rito religioso o sacrificale, sulla base della trentina di scheletri trovati nella "Fossa delle ossa" ad Atapuerca, insieme a un'ascia in quarzo rosa, ma potrebbe trattarsi di un seppellimento per impedire l'attacco di animali.
Caratteristiche
In generale, gli esemplari di H. heidelbergensis mostrano una continuazione delle tendenze evolutive che si sono presentate nel Pleistocene inferiore e medio.
Innanzitutto c'è un aumento delle dimensioni del cervello fino a raggiungere o superare i 1200 cc; la parte posteriore della volta cranica diventa più verticale; la calotta cranica è più allargata; la parete posteriore del cranio è verticale anziché arrotondata o inclinata; c'è una riduzione graduale di robustezza del cranico.
L'aumento del cervello può anche comportare un aumento nella sua complessità, anche se questo è difficile da determinare e può significare cambiamenti nel comportamento che conducono ad una maggiore abilità nel raggiungere facilmente le risorse nutrizionali, presupposto indispensabile per lo sviluppo del cervello.
L'area di Broca e le altre aree per il linguaggio sono ben sviluppate e i lobi frontali e parietali del cervello sono ingranditi.
La faccia è molto grande, corta e lievemente prognata, con un grande naso, la fronte meno sfuggente e toro sopraorbitale più piccolo di quello di H. erectus.
![Facial (A) and lateral (B) views of crania. Clockwise from top left: Homo erectus (replica, Sangiran, Java), heidelbergensis (Broken Hill, Zambia), sapiens (recent, Indonesia), and neanderthalensis (replica, La Ferrassie, France). All pictures © The Natural Histroy Museum London. [Color figure can be viewed in the online issue, which is available at wileyonlinelibrary.com.]](https://www.researchgate.net/profile/Christopher-Stringer/publication/227710322/figure/fig3/AS:280304112685064@1443841116689/Facial-A-and-lateral-B-views-of-crania-Clockwise-from-top-left-Homo-erectus.png)
Viste frontali (A) e laterali (B) del cranio. In senso orario da sinistra in alto:
Homo erectus
(replica, Sangiran, Java),
H. heidelbergensis
(Broken Hill, Zambia),
H. sapiens
(recente, Indonesia) e H.
neanderthalensis
(replica, La Ferrassie, Francia). (© The Natural Histroy Museum London - Clicca sull'immagine per i crediti)
La mandibola è molto robusta per l'attacco di forti muscoli masticatori, con i denti disposti a parabola, più piccoli degli Ominini precedenti, ma più grandi degli umani moderni e il mento è sfuggente.
Gli arti inferiori sono relativamente lunghi, simili a quelli di H. sapiens, in contrasto con quelli del successivo H. neanderthalensis.
La specie raggiunge un'altezza considerevole: i maschi erano alti oltre i 175 cm per 62 Kg, le femmine 160 m e 51 Kg. H. hidelbergensis è solitamente meno robusto di H. erectus, ma più degli esseri umani moderni.
Implicazioni evolutive
Le caratteristiche sia primitive sia derivate collegano H. heidelbergensis a H. neanderthalensis e a H. sapiens.
Come abbia visto a proposito di H. antecessor, per alcuni studiosi quest'ultimo sarebbe l'antenato diretto, perciò si sarebbe evoluto in Europa e una parte della popolazione sarebbe migrata in Africa.
Per altri l'antenato è H. ergaster che, evolutosi localmente in H. heidelbergensis, è uscito dall'Africa 800.000 anni fa (Out of Africa II), oppure la speciazione è avvenuta in Europa dopo l'uscita. Una parte della popolazione rimasta nel continente Africano - quella chiamata H. rhodesiensis - avrebbe poi originato H. sapiens.
Per quanto riguarda i discendenti, da esso deriva direttamente H. neanderthalensis e H. denisovensis nel primo caso, e anche H. sapiens nel secondo.
Le relazioni con l'Uomo di Denisova saranno meglio definite in quella sede.
Un fossile che sembra avere le giuste caratteristiche per essere alla base della divergenza neanderthal-sapiens è il "Cranio di Ceprano" (Homo cepranensis), datato 900/800mila anni fa, trovato a Ceprano, nel Lazio, perché non presenta ancora caratteri di H. neanderthalensis e corrisponde a un ancestrale H. heidelbergensis, simile al coevo H. erectus trovati in Algeria, e questo fa supporre che questi Ominini siano giunti in Italia dal Nord Africa.
Homo neanderthalensis
Homo neanderthalensis è un Ominino comparso circa 300.000 anni fa (Paleolitico medio) in Europa occidentale e centrale, espanso poi in Medio Oriente durante i rigidi inverni per raggiungere con pochi individui l'Asia (Monti Altaj), fino a scomparire 37/28.000 anni fa. In alcuni testi lo troviamo indicato anche Homo sapiens neanderthalensis, perché considerato sottospecie di H. sapiens.
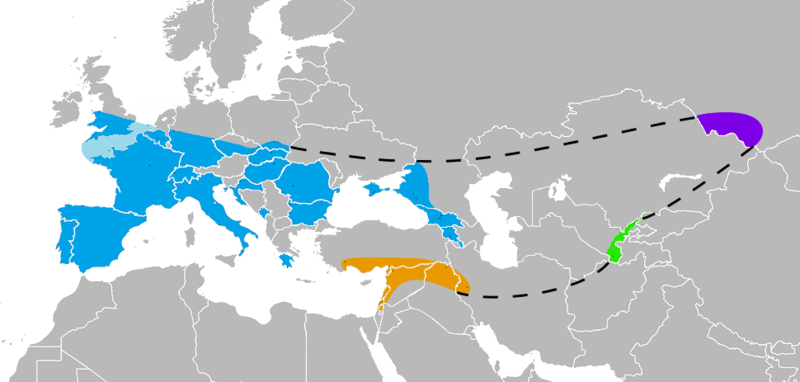
Estensione di
H. neanderthalensis
(Crediti:
Nilenbert, Nicolas Perrault III
- CC BY-SA 3.0)
{kind=link}
Il primo reperto è stato trovato nel 1829, ma solo nel 1856 è stato riconosciuto come umano un cranio rinvenuto nella valle di Neander vicino a Düsseldorf in Germania, dove "thal" in tedesco significa "valle" e da qui il nome H. neanderthalensis, "Uomo della valle di Neander" assegnato nel 1864. Da allora sono stati trovati moltissimi fossili di individui di tutte le età che hanno fornito un quadro completo delle caratteristiche anatomiche, fisiologiche e comportamentali.
In base all'età si possono individuare 3 periodi: Neandertaliani antichi (300.000 - 130.000 anni), trovati in Francia, Croazia e Saccopastore nel Lazio; Neandertaliani classici (130.000 - 45.000 anni), trovati nell'Europa occidentale, più robusti di quelli rinvenuti in altre zone; Neandertaliani tardivi (45.000 - 28.000), trovati in Portogallo e Spagna.



Ambiente e modo di vita
 H. neanderthalensis è stata la prima specie umana ad adattarsi e svilupparsi in ambienti freddi e con limitate risorse. Viveva, infatti, ai margini dei ghiacciai dell'Europa e parte dell'Asia in ambiente prevalentemente boscoso, ma si adattava anche alle regioni a clima temperato e mediterraneo. Seguendo il ritirarsi dei ghiacciai del Quaternario, è risalito a Nord fino all'Inghilterra, per poi scendere a Gibilterra, Altamura e Palestina quando il gelo riconquistava il terreno.
H. neanderthalensis è stata la prima specie umana ad adattarsi e svilupparsi in ambienti freddi e con limitate risorse. Viveva, infatti, ai margini dei ghiacciai dell'Europa e parte dell'Asia in ambiente prevalentemente boscoso, ma si adattava anche alle regioni a clima temperato e mediterraneo. Seguendo il ritirarsi dei ghiacciai del Quaternario, è risalito a Nord fino all'Inghilterra, per poi scendere a Gibilterra, Altamura e Palestina quando il gelo riconquistava il terreno.
Per difendersi dal freddo si copriva con le pelli avvolte intorno al corpo e si rifugiava nelle grotte, dove viveva in piccoli gruppi. Sempre con le pelli costruiva dei ripari estivi all'aperto.
Sapeva usare il fuoco, come testimoniano le tracce di focolari, con il quale si riscaldava, si difendeva e cuoceva i cibi.

Si nutriva di un'ampia gamma di alimenti, soprattutto legati all'area in cui viveva, con abbondante percentuale di carne. Se l'ambiente era boscoso, nella dieta entravano molte piante e piccoli animali; nelle aree costiere si nutriva dei prodotti ittici mentre nelle steppe e nelle pianure cacciava la megafauna (mammut, cervi, renne, rinoceronte lanoso, ecc.), entrando in competizione con leoni e lupi nel difficile territorio. Non è dato sapere se usasse armi da lancio, ma probabilmente si serviva della tecnica dell'imboscata.


Accanto ai resti sono stati trovati numerosi strumenti, molto più perfezionati della cultura precedente, appartenenti all'industria litica del Musteriano (Modo 3), con punte, raschiatoi, attrezzi denticolati, amigdale ben realizzate.

Punte Levallois del Musteriano al Museo di Tolosa (Crediti:
Didier Descouens
- CC BY-SA 4.0)
{kind=link}
Seppelliva i morti, a volte con armi, strumenti litici e cibo e anche fiori ed erbe medicinali, e ciò potrebbe far pensare che credesse a una vita dopo la morte, ma potrebbe anche essere stato fatto per evitare lo smembramento delle prede.

Ricostruzione della tomba di La Chapelle-aux-Saints al Musée de La Chapelle-aux-Saints (Crediti:
120 / V. Mourre
CC BY-SA 3.0)
{kind=link}
 La capacità simbolica e artistica è dimostrata dall'uso dell'ocra, da ornamenti fatti di conchiglie, penne di uccello o di osso. Anche se ancora controverse, sono da segnalare espressioni artistiche in grotte spagnole di cui abbiamo già accennato in precedenza.
La capacità simbolica e artistica è dimostrata dall'uso dell'ocra, da ornamenti fatti di conchiglie, penne di uccello o di osso. Anche se ancora controverse, sono da segnalare espressioni artistiche in grotte spagnole di cui abbiamo già accennato in precedenza.
È probabile che l'uomo di Neanderthal fosse in grado di parlare, esprimendosi anche con concetti abbastanza complessi, poiché non è ammissibile che il suo livello di vita, la concezione della comunità e della società, l'astrazione insita nelle credenze, potessero essere espressi senza la trasmissione verbale delle idee. Possedeva il gene FOX P2 nel genoma, ma con varianti e probabilmente non erano in grado di esprimere una gamma di suoni analoga a quella nostra.
Sono stati trovati individui che avevano subito malattie o ferite paralizzanti e la loro sopravvivenza può essere compresa solo all'interno di una vita sociale molto complessa, dove altri membri si occupavano del loro mantenimento e della cura, facendo uso di piante medicinali.
Le cure erano spesso necessarie perché traumi e lesioni di varia natura erano molto frequenti e perciò la vita media non andava oltre i 40 anni.
Caratteristiche
In passato H. neanderthalensis era comunemente immaginato come "l'uomo delle caverne", tozzo e curvo di spalle, rozzo, ottuso, peloso, di pelle scura, armato di clava, poco più di uno scimmione rispetto al bianco civilizzato, secondo la classica idea della progressione lineare dell'evoluzione. Ciò era dovuto sia ai pregiudizi dei ricercatori, sia al fatto che alcuni individui anziani presentavano artrite cronica. In realtà, i Neandertaliani erano piuttosto simili all'uomo moderno, forti e intelligenti, a parte alcune caratteristiche secondarie e con una cultura che ha loro permesso di sopravvivere in un ambiente ostile per oltre 200.000 anni. Anche se la comunità scientifica ha rivisto nel tempo le sue posizioni, altrettanto non è accaduto nella cultura popolare, come dimostrano i numerosi racconti e film ambientati nella preistoria, ma anche in alcuni francobolli.

Ricostruzioni dal 1875 al 1930 (clicca sull'immagine per vedere la galleria)



Ricostruzioni moderne di un bambino di 9 anni di
Tom Björklund
(CC BY-SA 4.0), di una donna (Crediti:
Bacon Cph
- CC BY 2.5), di un anziano (Crediti:
Neanderthal-Museum, Mettmann
- CC BY-SA 4.0) (clicca sull'immagine per ingrandire)
{kind=link}
{kind=link}
H. neanderthalensis un corpo tozzo, membra corte, indice di individui adattati a vivere in un clima particolarmente freddo.
Doveva essere alto mediamente 155 (F) - 165 (M) cm, più basso di H. sapiens, di peso intorno ai 50 kg (F) e 65 kg (M), con muscolatura molto potente.
Il cranio è ormai molto grande, con volume cerebrale di 1300 - 1600 cc, mediamente superiore al nostro - per la maggior massa corporea più che per l'encefalizzazione - allungato e schiacciato, con una fronte bassa e sfuggente e sviluppo di una protuberanza nella parte posteriore del cranio, lo chignon.
Le arcate sopracciliari sono molto robuste e formano un rilievo continuo che sovrasta entrambe le orbite, grandi e arrotondate.
La regione naso-mascellare è molto sviluppata, con aspetto quasi rigonfio; la faccia è piuttosto proiettata in avanti lasciando uno spazio dietro il terzo molare (spazio retromolare); il naso è ampio, non schiacciato e molto largo per umidificare e riscaldare l'aria fredda e secca.
La mandibola è priva di mento osseo o ne presenta un semplice abbozzo; i denti sono simili a quelli dell'uomo attuale, ma molto più grandi.
Le ossa degli arti sono simili alle nostre, ma più corte e sensibilmente più robuste.

(Crediti:
120
- CC BY 2.5)
La gabbia toracica è a forma di botte, con la colonna vertebrale curvata verso l'interno nella regione toracica per compensare la ridotta curvatura della regione lombare. Il bacino è più ampio rispetto ai moderni umani.
Il colore della pelle varia secondo la regione ed è stato individuato un gene per i capelli rossi e la pelle chiara.
(Crediti:
Thilo Parg
- CC BY-SA 4.0)
{kind=link}
Implicazioni evolutive
Il percorso evolutivo di H. neanderthalensis parte da H. ergaster, attraverso H. antecessor in Europa ha origine H. heidelbergensis, oppure in Africa da H. ergaster origina H. heidelbergensis che si trasferisce in Europa. Da un piccolo gruppo di H. heidelbergensis deriva direttamente H. neanderthalensis, circa 300.000 anni fa.
H. neanderthalensis è un vicolo cieco dell'evoluzione e da esso non deriva H. sapiens ma hanno un antenato in comune: H. heidelbergensis o H. antecessor. Le analisi genetiche indicano in 800.000 - 500.000 il momento di separazione della linea che porta a H. sapiens.
Poiché l'uomo di Neanderthal non appartiene alla linea del sapiens, non può essere considerato una sottospecie (H. sapiens neanderthalensis) perciò oggi si preferisce H. neanderthalensis mentre noi siamo semplicemente sapiens e non H. sapiens sapiens.
I rapporti con l'Uomo di Denisova e con H. sapiens saranno esaminati nel prossimo paragrafo.
Le analisi del DNA ci forniscono anche un altro dato: la presenza di una percentuale tra 1 e il 4% di DNA neandertaliano nelle moderne popolazioni europee, asiatiche e dei nativi americani, ma non in quelle dell'Africa subsahariana. Quest'ultimo dato è stato contestato da studi recenti di alcuni scienziati, che hanno ritrovato una certa percentuale di DNA neandertaliano anche nella popolazione africana.
Come si spiega la presenza di questo DNA se non siamo direttamente collegati dal punto di vista filogenetico?
Studi del 2006 e del 2010 indicano che tra H. Neanderthal e H. sapiens ci sono stati uno (improbabile) o più eventi di incrocio, l'ultimo dei quali si è verificato tra 80.000 e 50.000 anni fa. Poiché in quel periodo H. sapiens era già uscito dall'Africa e non ancora diffuso in Asia ed Europa e l'attuale popolazione africana è priva (forse) dei frammenti di DNA neandertaliano, l'ibridazione deve essere avvenuta nel Vicino Oriente dove le due specie convivevano.
Rimanendo nell'ipotesi dell'ibridazione, secondo un recente studio un primo incrocio non sarebbe avvenuto direttamente tra H neanderthalensis e H. sapiens, ma tra H. heidelbergensis e un'antica popolazione di H. erectus già presente in Eurasia, circa 600.000 anni fa, seguito da molti altri episodi di ibridazione.
Per giustificare la possibilità di ibridazione, occorre considerare che almeno alcune popolazioni fossero in grado di incrociarsi e quindi non erano vere specie biologiche in quel momento nel tempo e nello spazio geografico, oppure si deve parlare non di specie umane distinte, ma di popolazioni umane distinte, oppure ancora che l'incapacità di riprodursi non si applicasse alle specie recenti del genere Homo.
Altri studi negano invece che ci siano evidenze di incroci tra le due specie, anche se non è possibile affermare definitivamente che l'incrocio non abbia avuto luogo. Bisogna anche tenere conto che manca nei moderni umani il cromosoma Y e il DNA mitocondriale dei Neandertaliani e questo renderebbe sterili gli eventuali ibridi. L'assenza del DNA mitocondriale potrebbe però essere spiegata con incroci in cui il padre è un Neanderthal.
La presenza di DNA neandertaliano potrebbe allora essere spiegata come il residuo di un antenato comune, piuttosto che tramite l'ibridazione, ma anche questo viene contestato per l'assenza di questo DNA nelle popolazioni africane. Tuttavia, la recente smentita di questo dato potrebbe rivalutare tale ipotesi.
Altri ancora sostengono la possibilità che siano valide entrambi le ipotesi, perciò i frammenti genetici derivano sia dall'antenato comune sia da incroci.
Estinzione
La scomparsa di H. neanderthalensis avvenne in maniera piuttosto rapida e misteriosa circa 40/30.000 anni fa e altrettanto rapida fu l'apparizione di H. sapiens, che si diffuse molto velocemente nelle parti più lontane del continente asiatico, in Australia, in Europa e in America, sostituendo le specie esistenti, dopo un periodo di migliaia di anni di coesistenza in alcune aree.
Le ipotesi di estinzione formulate sono molteplici.
- Una delle prime ipotesi è lo scontro diretto tra le due specie - o addirittura cacciati - che ha spinto fuori dal loro habitat i Neandertaliani verso aree inospitali, fino a giungere all'estinzione, a un vero genocidio, tuttavia abbia appena affermato che in diversi luoghi sono vissuti insieme per molto tempo.
- Altra vecchia ipotesi vede in H. sapiens un'intelligenza superiore che si rifletteva in una migliore capacità di adattamento e di soluzioni dei problemi e situazioni in cui si veniva a trovare.
- La superiore intelligenza ha portato H. sapiens a renderlo un migliore cacciatore, a sviluppare una cultura complessa basata su tecniche avanzate di commercio, cosa che portava più tempo libero rispetto ad una cultura basata sulla caccia, da impiegare per sviluppare specializzazioni non strettamente legate alla sussistenza.
- Inoltre, le conoscenze tecnologiche, da sempre ritenute superiori in H. sapiens, li hanno dotati di migliori armi per la caccia, della capacità di vestirsi ecc. che hanno permesso di vivere in qualunque ambiente pur essendo meno adattati al clima freddo rispetto ai Neandertaliani.
- La cultura si manifestava anche nel pensiero simbolico, ritenuto praticamente assente nei Neandertaliani e nel linguaggio più complesso e articolato in H. sapiens. In realtà sono ben presenti anche nei Neandertaliani espressioni simboliche e, inoltre, l'assetto culturale non doveva essere molto diverso nelle sue specie, come si rileva dalle documentazioni archeologiche.
- Le malattie portate dall'H. sapiens sarebbero state fatali perché è mancato il tempo di sviluppare adeguate difese immunitarie all'interno di una popolazione con scarso numero di individui aventi bassa diversità genetica.
- Proprio la bassa variabilità genetica dovuta all'incrocio tra consanguinei all'interno di una piccola popolazione, avrebbe portato all'espressione di geni dannosi e, in ogni caso, l'esiguo numero espone costantemente la popolazione al rischio di estinzione, considerando anche che vi sono limitate possibilità di accoppiamento.
- Il rigido clima europeo avrebbe influito sul livello di fertilità, rendendolo inferiore a quello dei sapiens, condannandoli così a una lenta scomparsa. Infatti, gran parte dei resti di H. neanderthalensis appartiene a bambini e a individui che non arrivano a 40 anni.
- Circa 30 000 anni fa il clima è diventato più freddo e secco, trasformando le foreste in steppe e questo costrinse le popolazioni neandertaliane a migrare in ambienti con clima più mite. Il ritorno di una fase più calda ha ripopolato le precedenti aree con H. sapiens, che aveva una maggiore capacità di insediamento. Tuttavia sappiamo che per millenni H. neanderthalensis ha superato diverse fasi di oscillazione climatica.
- Il peggioramento del clima ha ridotto drasticamente la capacità dell'ambiente di dare sostentamento agli esseri viventi e da qui l'innescarsi di una competizione per le risorse in cui solo H. sapiens è risultato vincitore perché, rispetto a H. neanderthalensis, richiedeva un minore apporto di energia dal cibo.
- Una dieta differenziata, marcatamente onnivora, forse è stata una delle armi in più dell'H. sapiens, che si impose su H. neanderthalensis, il quale, invece, si cibava prevalentemente della megafauna che andava progressivamente estinguendosi. Sappiamo, tuttavia, che anche quest'ultimo aveva una dieta sufficientemente varia.
- Sempre nel campo ambientale, un'ipotesi sulla loro estinzione potrebbe essere dovuta a un'importante diminuzione del campo magnetico terrestre, avvenuta intorno a 40.000 anni fa e durata all'incirca 2.000 anni, con il conseguente aumento della quantità di raggi ultravioletti che giungevano sulla superficie terrestre. Essa avrebbe colpito maggiormente H. neanderthalensis con mutazioni negative a causa della sua minore variabilità genetica e alla mutazione di un recettore molto sensibile a questa radiazione.
- Una spiegazione controversa potrebbe essere quella secondo cui H. neanderthalensis si stava ibridando con H. sapiens per cui il suo genoma sarebbe stato inglobato e poi diluito in quest'ultimo perché presente con un maggiore numero di individui e ciò spiegherebbe anche la progressiva sostituzione, nel corso degli incroci tra femmine neandertaliane e maschi sapiens, del cromosoma Y neandertaliano con quello dei sapiens. Secondo alcuni autori, inoltre, l'incrocio tra femmine neandertaliane e maschi sapiens avrebbe generato prole sterile, oppure i maschi ibridi, per la scomparsa del cromosoma Y, avrebbero avuto una fertilità ridotta.
- Le donne Neanderthal aiutavano gli uomini nella caccia di bisonti, gazzelle, cervi e tutta la selvaggina che prosperava in Europa nell'era glaciale. Costretti ad affrontare gli animali molto da vicino, uomini, donne e bambini, erano esposti a gravi rischi, mentre con la suddivisione dei lavori tra i sessi in H. sapiens, si affidavano alle femmine ruoli stanziali e meno pesanti in modo da potersi prendere cura più facilmente della prole, aumentando le possibilità di sopravvivenza.
In realtà sembra ragionevole pensare che non esista una sola e unica spiegazione della scomparsa dei Neanderthal, ma probabilmente un insieme di spiegazioni, anche se H. sapiens può avere avuto un ruolo importante nell'estinzione di H. neanderthalensis.
Homo denisovensis
Homo denisovensis, se lo si considera come specie (non ancora definitivamente determinato), o Denisovan, è un Ominino i cui resti sono stati ritrovati nella grotta di Denisova nei Monti Altaj della Siberia (Russia) e da qui il nome specifico, che a sua volta deriva da un eremita russo, Denis, del diciottesimo secolo.
La grotta è stata, in vari periodi, abitata da tre specie di Ominini: i Denisova, H. neanderthalensis e H. sapiens.
La determinazione specifica, unico caso, è stata fatta geneticamente su una falange fossile di una giovane femmina di circa 41.000 anni fa (o, secondo nuove stime, 76 - 51.000 anni fa), ma i fossili sono stati ritrovati a partire dagli anni settanta.
Si stima che sia vissuto nel Paleolitico medio, durante l'ultima era glaciale, nell'Asia orientale, ma probabilmente viveva anche in Estremo Oriente, anche se al momento non sono state trovate prove fossili.
Le datazioni radiometriche indicano un'età della specie compresa tra 70.000 e 30.000 anni fa, ma in una grotta del Tibet, a circa 3.280 metri di altitudine, è stato trovato nel 1980 un frammento di mandibola che potrebbe appartenere all'Uomo di Denisova, datato 160.000 anni.

Un ritratto di una giovane donna Denisovan basato su un profilo ricostruito da mappe di metilazione del DNA (Crediti: Maayan Harel)
Ambiente e modo di vita
H. denisovensis è vissuto in un periodo in cui il clima, nella zona di Denisova era fresco e secco, con un paesaggio di foresta decidua rada o di tundra, che si alternava con fasi più calde con una vegetazione di pini e betulle. Il ritrovamento nella grotta carsica del Tibet, invece, mostra un adattamento alle alte quote. Questo tipo di ambiente non era il principale o esclusivo perché, nei momenti in cui il livello marino era basso, si aprivano passaggi verso le calde aree delle isole dell'Indonesia.
Nella grotta sono stati trovati moltissimi strumenti e monili che coprono tutte le fasi del Paleolitico prodotti da H. neanderthalensis e H. sapiens, ma nessuno è direttamente associabile all'Uomo di Denisova, anche se è presumibile che possedessero lo stesso livello culturale delle altre popolazioni coeve.
Caratteristiche
Di H. denisovensis sono stati trovati pochissimi fossili: l'osso di un mignolo appartenente a una ragazza di 7 - 13 anni, tre denti, alcuni frammenti di ossa lunghe, un frammento di osso parietale, più un frammento di mandibola nella grotta del Tibet.
Per questo motivo la possibile ricostruzione delle caratteristiche fisiche deriva dalla sola analisi del DNA nucleare e mitocondriale studiando la metilazione, uno dei controlli dell'espressione genica che modifica il fenotipo senza alterarne il genotipo, in modo da ottenere una previsione sull'aspetto fisico. Ne risulta che i Denisoviani avevano alcuni caratteri in comune con H. neanderthalensis, altri con H. sapiens, altri ancora erano unici.
Il frammento della mandibola tibetana, che aveva ancora due denti, presenta caratteristiche arcaiche: è robusta, con arcata dentale ampia e mento sfuggente, i molari sono molto grandi e privi delle caratteristiche specifiche dei Neandertaliani e non assomigliano nemmeno a quelli dell'uomo moderno, ma sono più vicini, piuttosto, a quelli di H. erectus o addirittura a quelli dell'Australopiteco.

Molare di Denisova 4 (Crediti:
Thilo Parg
- CC BY-SA 4.0)
{kind=link}
I dati emersi dal genoma della ragazza indicano la presenza di 56 caratteristiche anatomiche, di cui 34 relative al cranio, che differenziano H. denisovensis da H. neanderthalensis e da H. sapiens.
Il cranio è allungato e appiattito, con la fronte bassa, il volto ampio e sporgente, il naso grande, caratteristiche presenti nei Neandertaliani, ma con un'espansione laterale propria della specie.
Anche il bacino e la gabbia toracica sono ampi e assomigliano a quelli dei Neandertaliani, ma le mani dovevano essere più simili a quelle dell'uomo moderno.
La ragazza era di pelle scura, con gli occhi e i capelli castani e aveva la capacità di adattamento alle alte quote.
Complessivamente questi Ominini dovevano essere molto robusti.
Implicazioni evolutive
Le analisi genetiche di cui abbiamo accennato e quelle fatte su un fossile di H. heidelbergensis, suggeriscono che quest'ultimo sia l'antenato comune di H. denisovensis, H. neanderthalensis e, con le precisazioni fatte in precedenza, anche di H. sapiens.

Circa 1,9 milioni di anni fa H. ergaster, o forse prima H. habilis, è uscito dall'Africa espandendosi verso oriente e, giunto in Asia, si è evoluto in H. erectus.
Tra 800.000 e 600.000 anni fa un'altra popolazione di H. ergaster, evolutasi in H. heidelbergensis, è uscita dall'Africa (Out of Africa II) espandendosi nell'Eurasia e qui circa 400.000 o forse 600.000 milioni di anni fa si divise in due gruppi. Il gruppo che si è espanso a Occidente ha originato H. neanderthalensis, che in seguito si è spostato anche a Est, mentre il gruppo che è andato a Oriente si è evoluto in H. denisovensis. La separazione potrebbe essere stata causata dall'espansione verso sud della calotta glaciale, che ha creato una barriera fisica. Infine, una popolazione di H. heidelbergensis rimasta in Africa ha dato i natali a H. sapiens, che successivamente è giunto in Asia tra i 60.000 e i 40.000 anni fa, dove ha incontrato gli altri due Ominini.

(Crediti:
John D. Croft
- CC BY-SA 3.0)
{kind=link}
Attraverso lo studio del DNA si è scoperto che H. denisovensis, H. neanderthalensis e H. sapiens si sono incrociati tra loro.
Un incrocio molto antico sarebbe avvenuto 700.000 anni fa tra l'antenato dei Denisoviani e Neandertaliani, cioè H. heidelbergensis, e una specie molto arcaica uscita dall'Africa, forse H. erectus.
Dopo la separazione tra Denisoviani e Neandertaliani, quest'ultimo si è incrociato con H. sapiens e di questo ne abbiamo già parlato in precedenza.
Anche le popolazioni di H. neanderthalensis recenti, provenienti dalla Croazia, e H. denisovensis, si sono incrociate più volte nell'area della Siberia. Ne abbiamo prova in una ragazza di almeno 13 anni, vissuta circa 90.000 anni fa a Denisova, che è l'ibrido tra una madre di H. neanderthalensis e il padre H. denisovensis e, sempre nella grotta di Denisova, il 17% del genoma denisoviano deriverebbe da incroci con Neandertaliani.
Vediamo ora le relazioni con l'uomo moderno.
Nella grotta di Denisova sappiamo che, oltre ai Neandertaliani e Denisoviani - che si sono sicuramente incrociati -, vivevano anche i sapiens, di cui non abbiamo prove di ibridazione. Le attuali popolazioni asiatiche e i nativi americani, tuttavia, hanno ricevuto dai Denisoviani lo 0,2% del patrimonio genetico, forse residuo di precedenti incroci, ma i Melanesiani e gli indigeni Australiani ne possiedono una parte più cospicua, il 5-6%, dimostrando che si sono incrociati più volte.
In particolare, un gruppo di Denisoviani del Nord si è incrociato con H. sapiens nell'Asia orientale portando ai moderni asiatici dell'est (Cinesi e giapponesi) una piccola percentuale di DNA denisoviano, mentre una seconda popolazione, più meridionale, di cui non si conoscono reperti fossili, dopo una serie di ibridazioni nel sud-est asiatico, ha lasciato quelle importanti tracce di DNA che ritroviamo nei moderni Melanesiani, negli aborigeni Australiani e in tutta l'area indonesiana. In queste zone, una prima ibridazione sarebbe avvenuta circa 46.000 anni fa, mentre un'altra attorno ai 30.000 anni fa, ma forse fino a 15.000 anni fa.
Le popolazioni tibetane, inoltre, avrebbero ereditato i geni in grado di sopportare la carenza di ossigeno nella catena Himalayana da un gruppo di Denisoviani che si era stabilito in Tibet e poi incrociati con H. sapiens circa 30.000 - 40.000 anni fa.
Homo floresiensis
I resti di una quindicina di Ominini, che si sono conservati in uno stato di non completa fossilizzazione, sono stati ritrovati dal 2003 nelle profondità di una grotta a Liang Bua, nell'isola di Flores in Indonesia, da un gruppo di archeologi australiani e indonesiani. A essi è stato assegnato il nome Homo floresiensis dalla località di ritrovamento, tuttavia persistono ancora studiosi che ritengono non si tratti di una nuova specie, ma di esemplari di H. sapiens affetti da una qualche patologia come microcefalia, sindrome di Down o cretinismo. Per la statura incredibilmente bassa (poco più di un metro) e il piccolo cervello, gli è stato assegnato il soprannome "Hobbit" in riferimento al popolo descritto nella saga di J.R.R. Tolkien. Il cranio completo di una donna di circa 30 anni funge da olotipo.
La prima datazione fornita era di 18.000 anni, ma nuove datazioni radiometriche, effettuate sui sedimenti e sugli strumenti litici rinvenuti nella grotta, risalenti a 190.000 - 50.000 anni fa, suggeriscono un'età più antica per questo Ominino: 100.000 - 60.000 anni.
Nella stessa isola, nel sito di Mata Menge, 70 chilometri ad est da Liang Bua, sono stati trovati i resti di una mandibola e sei denti appartenenti ad almeno tre individui diversi, solo che sono vecchi di 700.000 anni, insieme a moltissimi strumenti litici datati a quasi un milione di anni fa. Le caratteristiche suggeriscono che potrebbero essere della stessa specie, forse più primitivi. Se confermato, viene escluso in maniera definitiva che si tratti di H. sapiens, arrivato in quelle aree 60.000 anni fa.
Curioso il fatto che sull'isola si trova il villaggio di Rampasasa i cui abitanti hanno un'altezza media di 145 cm. Le analisi genetiche indicano, tuttavia, che non si tratta dei discenditi di H. floresiensis, ma un particolare adattamento genetico.

(Crediti:
Cicero Moraes
et alii -
http://arc-team-open-research.blogspot.it/2015/08/homo-floresiensis.html
- CC BY 4.0)
{kind=link}
Ambiente e modo di vita
 L'isola di Flores era, come lo è ancora, una zona di foresta tropicale, ricca di molte specie vegetali diverse, ma con bassa quantità per ciascuna, soprattutto di quelle alimentari, perciò le risorse per l'alimentazione non erano abbondanti. Questo spiega il nanismo insulare, onde sfruttare al meglio le scarse risorse.
L'isola di Flores era, come lo è ancora, una zona di foresta tropicale, ricca di molte specie vegetali diverse, ma con bassa quantità per ciascuna, soprattutto di quelle alimentari, perciò le risorse per l'alimentazione non erano abbondanti. Questo spiega il nanismo insulare, onde sfruttare al meglio le scarse risorse.
Nell'area sono stati trovati resti di numerosi animali nani del genere Stegodon, antenato dell'elefante moderno con tracce di macellazione, draghi di Komodo, una lucertola gigante, ratti, cicogne giganti, avvoltoi, pipistrelli, probabilmente alimenti di questi Ominini.
Ossa bruciate, roccia spaccata dal fuoco e un possibile focolare costituito da un cerchio di rocce alterate, mostrano che hanno fatto uso del fuoco.
In associazione sono stati trovati strumenti litici, piccoli come lo erano i loro artefici, ma qualche studioso ritiene che debbano essere attribuiti a H. erectus vissuto sull'isola.
Caratteristiche
H. floresiensis presenta un miscuglio di caratteristiche primitive e moderne, con in più alcune caratteristiche del tutto particolari, come la sua ridotta statura.
Era alto circa 106 cm (sulla base dello scheletro femminile ritrovato) e pesava 25 - 30 kg. La capacità cranica era di soli 380 cc, inferiore a quella di tutti gli Ominini conosciuti, con una cresta sopracciliare divisa in due sezioni, fronte sfuggente e assenza di mento che lo avvicinano a H. erectus. La riduzione del cervello non sembra avere influito sulle capacità cognitive, come dimostrano le abilità tecnico-culturali citate sopra.

(Crediti:
Rama
and
one more author
- CC BY-SA 3.0 fr)
{kind=link}
I denti complessivamente sono piccoli, ma grandi in proporzione alle dimensioni del corpo. I canini e i premolari hanno una morfologia primitiva, mentre i molari sono moderni.
L'anca e gli arti sono robusti e simili a quelli dell'Australopiteco, soprattutto per quanto riguarda la spalla, alzata in avanti, e il polso.
I piedi sono molto lunghi in rapporto agli arti inferiori, con proporzioni più simili a quelle delle antropomorfe africane piuttosto che a quelle umane. L'alluce, pur se allineato alle altre dita come negli umani, è molto più corto di esse, una caratteristica decisamente primitiva. Infine, il piede è piatto, non leggermente arcuato come il nostro.

(Crediti:
Emőke Dénes
- CC BY-SA 4.0)
{kind=link}
Implicazioni evolutive
Si diceva sopra che alcuni studiosi ritengono che H. floresiensis sia in realtà H. sapiens affetto da una qualche patologia o disturbo della crescita, come il nanismo, la microcefalia, la sindrome di Down. Se guardiamo però le date, H. floresiensis era già presente sull'isola quando è arrivato H. sapiens, perciò l'ipotesi è da escludere. Inoltre, la microcefalia o la sindrome di Down non comportano anche una modifica nella proporzione degli arti, tanto da far loro assumere un aspetto scimmiesco.
Molto quotata è anche l'ipotesi che derivi da H. erectus - soprattutto per la morfologia dei denti - il quale, rimasto isolato per centinaia di migliaia di anni nell'isola, si sarebbe evoluto nella nuova specie, con dimensioni ridotte, come adattamento alle condizioni dell'isola, dove una dieta con poche calorie e la mancanza di grossi predatori avrebbe avvantaggiato le caratteristiche fisiche piccole (nanismo insulare). La statura molto bassa indica che anche gli uomini sono soggetti alle stesse forze evolutive di altri mammiferi che si rimpiccioliscono fino a raggiungere dimensioni nane quando rimangono isolati o sotto la pressione dell'ambiente, come è successo all'elefante nano del genere Stegodon. In altri casi, per difendersi dai predatori, alcuni animali diventano molto più grandi come il grosso roditore o la cicogna alta 182 cm.
Ma anche questa ipotesi, secondo recenti studi, sarebbe da scartare perché possiedono caratteristiche così primitive nella forma del cranio e nei grandi piedi da far ritenere che discenda da un Ominino africano molto antico, forse H. habilis, H. rudolfensis o ancora precedente (Australopithecus che però non è mai stato trovato fuori dall'Africa). Secondo alcuni autori, infatti, H. habilis e H floresiensis potrebbero avere un antenato africano in comune da cui H. floresiensis si sarebbe separato prima di H. habilis e quindi, evolutosi in Africa, si è spostato fino in Indonesia prima di qualsiasi altro Ominino. Potrebbe anche essere stato l'antenato comune a migrare e poi si sarebbe evoluto in H. floresiensis da cui prima si era separato H. habilis.

Come giunse in quella lontana isola H. floresiensis, che si trova a Est della Linea di Wallace, la linea che segna una sorta di barriera naturale alla migrazioni di diverse specie animali e vegetali? A piedi, fino a un certo punto, ma poi c'era un mare infido da attraversare (per quanto basso durante le glaciazioni) e quindi, nonostante l'opinione che solo H. sapiens fosse in grado di farlo, sembra proprio che sia stato lui il primo a giungervi con una qualche specie di zattera.
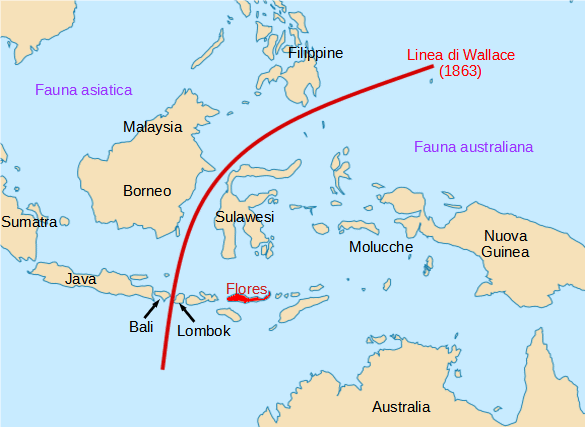
L'estinzione di H. floresiensis potrebbe essere stata causata dall'arrivo sull'isola di H. sapiens, con cui è entrato in competizione, perdendo la battaglia. Secondo alcuni autori, potrebbe essere stata anche un'eruzione vulcanica, che ha fatto estinguere anche gran parte della fauna che costituiva la sua fonte di sostentamento.
Homo luzonensis
Homo luzonensis è un Ominino del Pleistocene superiore trovato nella Grotta di Callao, nella regione di Peñablanca, sulla parte settentrionale dell'isola di Luzon (Filippine). Il nome specifico deriva dalla località di ritrovamento.
Il primo reperto, il terzo metatarso, è stato recuperato nel 2007 e poi, dal 2011 al 2015, un gruppo di scienziati coordinati dal paleontologo Florent Détroit ha trovato altri dodici fossili appartenenti a due adulti e un bambino: un femore del bambino, sette denti tra molari e premolari, due falangi dei piedi e due della mano.
La datazione radiometrica indica un'età di 67.000 anni, perciò si pensa che sia vissuto tra 50.000 e 67.000 fa, contemporaneamente a H. floresiensis, H. neanderthalensis, H. denisovensis e H. sapiens, ma presenta caratteristiche diverse da questi, giustificando l'assegnazione di una nuova specie. Nella Cagayan Valley, adiacente alla Grotta di Callao, sono stati trovati strumenti litici e ossa di un rinoceronte macellato di almeno 700.000 anni, ma non ci sono reperti che indichino chi possano essere gli artefici. Anche a Flores abbiamo visto che sono stati trovati strumenti della stessa epoca.

Grotta Callao nelle Filippine (
© Chad M/Flickr CC BY-NC-ND 2.0
)
Ambiente e modo di vita
H. luzonensis viveva in un ambiente boscoso, dove poteva agilmente arrampicarsi sugli alberi e probabilmente era un cacciatore di cervi e rinoceronti.
Caratteristiche
In base alle dimensioni del metatarso e dei denti, si presume che H. luzonensis fosse di corporatura gracile, di piccola statura, ma non quanto H. floresiensis.
Questo Ominino presentava un miscuglio di caratteristiche primitive e derivate, in una combinazione diversa rispetto agli altri Ominini suoi contemporanei.
Le caratteristiche primitive, simili a quelle di Australopithecus, si evidenziano nelle mani e nei piedi, mentre quelle derivate, con somiglianze a H. sapiens, le vediamo nei denti.
I denti sono complessivamente piccoli, ma i premolari sono piuttosto primitivi perché abbastanza grandi e con una terza radice, come in Australopithecus e H. erectus, mentre i molari sono simili a quelli dell'uomo moderno e molto piccoli.

(Crediti: Callao Cave Archaeology Project)
Le mani e i piedi sono diversi da quelle degli umani moderni perché le dita sono curve come quelle dell'Australopiteco e indicano una grande abilità nell'arrampicarsi sugli alberi. Gli scopritori si sono chiesti se si tratti di un adattamento al particolare ambiente o se sono stati ereditati da un antenato.
Implicazioni evolutive
Anche per H. luzonensis si pongono gli stessi interrogativi di H. floresiensis. Per alcuni autori deriverebbe dal più grande H. erectus che, soggetto al fenomeno del nanismo insulare, ha assunto nel tempo piccole dimensioni come adattamento alle limitate risorse disponibili.
Per altri, invece, sarebbe l'evoluzione di un'antica specie uscita dall'Africa prima della migrazione/espansione di H. ergaster. Le migrazioni sarebbero dunque 4 e non 3 e questo risolverebbe anche il problema della presenza di strumenti di 2,1 milioni di anni scoperti in Cina.
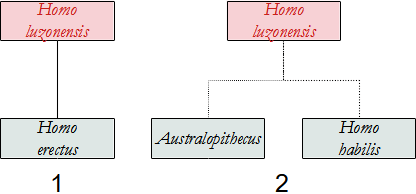
Ci chiede ancora una volta come abbia fatto a giungere in quest'isola, circondata da acque profonde, che non è mai stata collegata alla terraferma, così come è successo per l'isola di Flores, che dista 2000 km: zattere o evento fortuito, come uno tsunami? Sarebbe assai singolare un caso accidentale che si è ripetuto tre volte: Luzon, Flores, Sulawesi, dove anche qui sono stati trovati utensili ma non Ominini.

La loro estinzione, anche in questo caso, potrebbe dipendere dall'arrivo di H. sapiens.
Un'ultima domanda che attende risposta: H. floresiensis e H. luzonensis sono filogeneticamente collegati tramite un processo di speciazione allopatrica di un'originaria popolazione o si tratta in entrambi i casi di nanismo insulare?
Homo naledi
Homo naledi è un Ominino del tardo Pleistocene medio scoperto tra il 2013 e il 2014 nella provincia di Gauteng in Sudafrica, a 50 km nord-ovest di Johannesburg e a poca distanza dai siti di Sterkfontein e Swartkrans, dove è stato trovato Australopithecus africanus, da un'équipe di ricercatori diretta da Lee Berger, all'interno di una delle grotte carsiche del complesso Rising Star Cave (Grotta della Stella Nascente) chiamata Dinaledi Chamber. In una seconda grotta, a circa 100 metri dalla precedente, la Lesedi Chamber, sono stati trovati altri reperti. Le camere sono raggiungibili da uno stretto pozzo di 12 m e si trovano a circa 30 m di profondità.
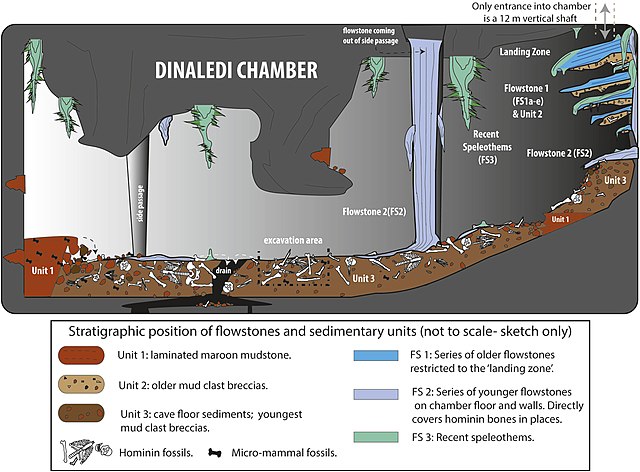
Dinaledi Chamber
(Crediti:
Paul HGM Dirks,
et al
.
- CC BY 4.0)
{kind=link}
Il nome specifico, annunciato nel 2015, significa "stella che sorge" nella lingua Sotho, in riferimento alla camera di ritrovamento e quindi "Uomo della stella nascente".
La prima camera conteneva circa 1550 resti non mineralizzati di almeno 15 individui di diversa età e sesso, mentre nella seconda sono stati ritrovati 131 reperti di almeno 3 individui, tra cui un maschio soprannominato "Neo", che nella lingua locale di Sesotho significa "dono", eccezionalmente completo, più di Lucy.
In un primo momento, viste alcune caratteristiche molto primitive, si pensava avesse tra 1 e 2 milioni di anni, ma nuove indagini mediante diversi metodi (luminescenza otticamente stimolata, serie dell'uranio, paleomagnetismo, risonanza di spin elettronico) hanno restituito un'età compresa tra 335.000 e 236.000 anni fa e questo significa che viveva contemporaneamente al H. heidelbergensis e/o ai primi H. sapiens.
Ambiente e modo di vita
L'ambiente è analogo a quello già descritto in A. sediba e P. robustus, con presenza di bosco dove questi Ominini prelevavano il cibo e si riparavano durante la notte, alternato a radure e praterie. Non è possibile risalire ad altre notizie perché sono stati trovati pochi resti di animali e piante. Le grotte non erano il loro rifugio perché al loro interno non sono stati trovati strumenti litici - ma potrebbero comunque essere stati in grado di fabbricarli - né tracce di fuoco, ma è probabile che ne conoscessero l'uso perché è necessario per entrare nelle grotte.
Dalle caratteristiche dei denti si desume che si nutrissero di vegetali che non richiedevano una grande masticazione e di alimenti in precedenza processati ma, contenendo residui sabbiosi, ne hanno abraso lo smalto. Probabilmente occupavano una nicchia ecologica diversa dagli Ominini contemporanei.

(Crediti:
Cicero Moraes et al.
- CC BY 4.0)
{kind=link}
Un mistero non ancora risolto è quello dell'accumulo di tanti corpi in un luogo difficilmente accessibile, che risultano essere deposti in un lungo arco di tempo.
Non ci sono tracce di predazione sui resti, perciò è improbabile che siano stati trascinati o fatti cedere all'interno da grossi carnivori.
Mancano prove di un loro trascinamento provocato dall'acqua, anche se in passato forse c'erano altri accessi da dove poteva entrare, ma in questo caso dovevano essere trascinati anche degli animali e detriti riferibili a un'alluvione.
Sempre per la mancanza di animali è improbabile che siano accidentalmente caduti da un'apertura carsica.
Sono forse entrati e poi rimasti intrappolati? Difficile accesso e troppi individui rendono improbabile quest'ipotesi.
Si è parlato di soppressione etnica da parte di un altro Ominino o di un'altra tribù - sarebbero delle specie di foibe - ma non ci sono tracce di violenza, oppure potrebbero essere stati imprigionati.
L'alternativa al momento più plausibile è una deposizione volontaria dei corpi, data la difficile accessibilità del luogo - ma non è detto che fosse sempre stato così -. Se si tratta di un rituale, non sarebbero più soltanto i Neandertaliani e i sapiens a mostrare un tale comportamento, tuttavia manca qualsiasi forma di simbolismo che lo provi (ma avevano la capacità di pensiero astratto?), inoltre, i reperti hanno una disposizione non solo orizzontale, ma anche verticale, con disarticolazione di alcuni elementi. Potrebbero invece averli depositati semplicemente per evitare che la decomposizione attirasse i predatori.
Caratteristiche
La grandissima quantità di reperti ci consente di avere un quadro completo dell'anatomia di H. naledi ma, diversamente da altri Ominini recenti, non è ancora sto possibile estrarre il DNA perché troppo degradato.
Come abbiamo visto per tanti Ominini, anche questo presenta un mosaico anatomico che lo avvicina al moderno H. sapiens, ma anche al lontano Australopithecus, soprattutto per quanto riguarda le dimensioni del cervello, la parte inferiore della gabbia toracica, il bacino e le falangi ricurve.
H. naledi maschio era alto mediamente 145 cm con un peso di 40 - 55 kg, un po' meno le femmine, perciò c'era un basso dimorfismo sessuale.
La morfologia del cranio assomiglia a quella di H. habilis o H. ergaster, con la parte posteriore flessa come nei moderni umani e la parte superiore simile a quella di H. erectus per la presenza di un ispessimento mediano (chiglia). La faccia è piatta, ma presenta un toro sopraorbitario ben sviluppato.

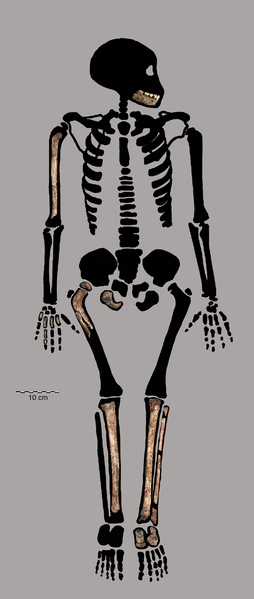 Il cervello ha le dimensioni medie di 560 cc (M) - 465 cc (F), come un Australopiteco, ma la struttura è simile a quella umana, soprattutto per quanto riguarda i lobi frontali.
Il cervello ha le dimensioni medie di 560 cc (M) - 465 cc (F), come un Australopiteco, ma la struttura è simile a quella umana, soprattutto per quanto riguarda i lobi frontali.
La mandibola e la mascella sono moderne, gracili, a forma di parabola, con denti complessivamente piccoli, ma il terzo molare è abbastanza grande. Gli incisivi e i canini hanno dimensioni analoghe a quelli di Australopitechi e Parantropi.
L'articolazione della spalla è elevata e facilita la sospensione, il tronco si allarga in basso e il bacino assomiglia a quello di un Australopiteco.
Le mani, i polsi e il pollice sono simili a quelle dell'uomo moderno, ma con falangi prossimali lunghe e ricurve, adatte per arrampicarsi, pur mantenendo una buona abilità manipolatoria, dato il pollice robusto.
Le gambe sono lunghe e sottili, ma con il collo del femore lungo e schiacciato antero-posteriormente come quello di Australopiteco.
Il piede è molto moderno, con arco longitudinale un po' ridotto, falangi prossimali leggermente curve, atto per la locomozione bipede in posizione eretta e per lunghe distanze.
(Crediti: Debra R. Bolter, Marina C. Elliott, John Hawks, Lee R. Berger - Bolter, D. R. (2020) - CC BY-SA 4.0)
Implicazioni evolutive
L'assenza di DNA, i tratti anatomici primitivi associati a una datazione recente rendono assai difficile formulare ipotesi sul percorso evolutivo di H. naledi. Ne citiamo alcune.
- Discende da una qualche forma di Australopiteco e, come si legge in alcuni giornali, sarebbe l'anello mancante tra questo e il genere Homo.
- Si tratta di un incrocio tra una specie di Homo con una qualche forma di Australopiteco.
- Deriva da una delle prime forme del genere Homo, come H. habilis o H. rudolfensis, quindi inizia il percorso 2 milioni di anni fa e poi rimane immutato per un lunghissimo periodo.
- Deriva da un ramo di un Ominino più recente e moderno, come H. ergaster.
- Ha un'origine ancora più recente (da H. antecessor/heidelbergensis?), un ramo parallelo a H. sapiens, che non ha dato germogli, un vicolo cieco dell'evoluzione.
H. sapiens potrebbe aver contribuito alla sua scomparsa?
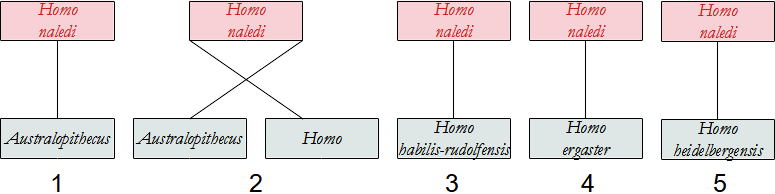
Homo sapiens
Homo sapiens è l'unica specie di Ominini esistente oggi sul nostro pianeta. Il nome specifico "sapiens" deriva dal fatto che l'uomo moderno si ritiene più sapiente, più intelligente, più abile delle altre specie che lo hanno preceduto e che con lui hanno convissuto.
Emblematico è il sottostante foglietto della Cambogia (2001!).

 Il Dna antico ha dimostrato che i sapiens che occupavano l'Europa avevano la pelle scura, più di quella dei Neandertaliani, che non andavano certo in giro con la clava in mano. "Cheddar man", il più antico fossile umano trovato in Gran Bretagna e risalente a 9.100 anni fa era nero, aveva capelli scuri ricci o ondulati e occhi azzurri o verdi. La carnagione chiara, contrariamente al pensare comune, è un'acquisizione recente in Europa, mentre era già presente da decine di migliaia di anni in alcuni Neandertaliani.
Il Dna antico ha dimostrato che i sapiens che occupavano l'Europa avevano la pelle scura, più di quella dei Neandertaliani, che non andavano certo in giro con la clava in mano. "Cheddar man", il più antico fossile umano trovato in Gran Bretagna e risalente a 9.100 anni fa era nero, aveva capelli scuri ricci o ondulati e occhi azzurri o verdi. La carnagione chiara, contrariamente al pensare comune, è un'acquisizione recente in Europa, mentre era già presente da decine di migliaia di anni in alcuni Neandertaliani.
Anche il volto e il cranio di H. sapiens conservano caratteristiche già presenti nei nostri antenati, mentre quelle di H. neanderthalensis, come il grande naso e il cranio basso sono derivate, acquisite in un epoca recente.
Descritto per la prima volta da Linneo nel 1758, non ha un olotipo di riferimento, a differenze delle altre specie animali.
Attualmente esiste una sola specie e non ci sono sottospecie o razze. Il Progetto Genoma Umano ha dimostrato la sostanziale uguaglianza dal punto biologico e genetico di tutti gli umani viventi e che dunque le razze non esistono: ci sono più differenze a livello genetico tra individui appartenenti alla stessa popolazione rispetto a due popolazioni diverse messe a confronto. Ciò è spiegabile con un'origine molto recente, unica e africana dell'attuale popolazione umana e perciò non c'è stato il tempo, né la presenza di barriere, per la formazione delle razze. Non solo, tra le popolazioni si è mantenuto un flusso genico e sembra anche che ci sia stato ad un certo punto della storia evolutiva un collo di bottiglia che ha sensibilmente ridotto la variabilità genetica. La differenza tra umani è quindi prevalentemente di tipo individuale e questo perché siamo tutti parenti (Eva mitocondriale e Adamo Y-cromosomiale), tutti di origine africana (Out of Africa) e tutti singolarmente diversi, perciò non esistono motivazioni scientifiche per giustificare l'esistenza della razze.
Secondo la maggior parte dei paleoantropologi, H. sapiens ha origine in Africa orientale circa 200.000 anni fa, ma un ritrovamento in Marocco, datato 300.000 anni, anticipa la data di comparsa, perciò in quel periodo doveva essere diffuso in tutto il continente. Qualche autore indica, invece, l'Africa settentrionale o il Botswana (Africa meridionale) come luogo di origine, ma gli studi non hanno convinto la comunità scientifica. Piuttosto, si può pensare al multiregionalismo africano.
Una recente scoperta di un reperto nel Peloponneso, datato 210.000 anni, congruente con la nuova datazione della sua comparsa, suggerisce che la prima migrazione (Out of Africa III) sia avvenuta molto presto e sia stata diretta verso l'Asia. In Medio Oriente ci sono siti che risalgono a 120.000 anni fa. Il ponte di terra araba probabilmente fungeva da corridoio per il Sud-est asiatico, proprio come aveva fatto per l'antenato H. ergaster. Secondo alcuni studiosi, questa migrazione sarebbe avvenuta con un numero ridottissimo di individui a causa di un collo di bottiglia, forse l'eruzione del supervulcano indonesiano Toba, che avrebbe raffreddato il clima rendendo ancora più difficile la sopravvivenza Una prova dell'eseguo numero di umani presenti, sarebbe fornita dalla scarsa variabilità genetica che la nostra specie esibisce rispetto ad altri animali.
In questa fase H. sapiens condivide il territorio si incontra con le altre popolazioni già presenti, con le quali convive, si ibrida o le assimila. Probabilmente l'impossibilità di incrociarsi con altre specie non si applica molto bene alle specie umane del genere Homo.
Tra 120.000 e 65.000 anni fa H. sapiens sarebbe uscito dall'Africa un'altra volta per cominciare la colonizzazione degli altri continenti. In questo caso non abbiamo più un'espansione a piccoli passi, dove a ogni generazione avanzava solo di pochi chilometri, ma è una vera migrazione, veloce, intenzionale, simile a quelle contemporanee, grazie ad una maggiore organizzazione sociale, un linguaggio articolato e a nuovi comportamenti simbolici, per cui le specie già presenti progressivamente scompaiono.
Rapidamente H. sapiens raggiunge tutta l'Asia centrale e meridionale e poi l'Asia orientale. In Mongolia e in Giappone abbiamo siti datati 50.000 anni.
Dall'Asia avrebbe raggiunto l'Australia circa 65.000 anni fa - ma ci sono segni di arte rupestre di 75.000 anni fa -, un territorio mai raggiunto da H. erectus, che dà origine ai nativi della Australia, della nuova Guinea, della Tasmania, mentre una popolazione sarebbe ritornata in Africa, come dimostra la presenza di DNA neandertaliano trovata nella popolazione africana.
Un'ondata, riferibile a 44.000 anni fa, è partita dal Medio Oriente verso l'Europa, dove le popolazioni prendono il nome di Cro-magnon. Anche in questo caso, studi condotti da un team internazionale coordinato da scienziati del Max Planck Institute for Evolutionary Anthropology di Lipsia (Germania), di cui fa parte anche l'Università di Bologna, su reperti trovati nel 2015 nella grotta di Bacho Kiro, a nord della Bulgaria, indicherebbe la presenza di H. sapiens in un intervallo tra 45.820 e 43.650 anni fa, e forse già 46.940 anni fa. Questo anticiperebbe la sua presenza in Europa di 2.000 anni e porterebbe a una compresenza con H. neanderthalensis di almeno 5.000 anni. Tra i reperti ivi trovati, ci sono oggetti in osso, simili a quelli che i Neandertaliani hanno prodotto molto più tardi in Francia, che testimonierebbero contatti e scambi culturali tra le due specie.
Anche gli strumenti litici trovati nella grotta di Lapa do Picareiro, in Portogallo, datati tra 41.000 e 38.000 anni fa, all'epoca della glaciazione Würm 2, suggeriscono che H. sapiens era presente nella regione 5000 anni prima di quanto si pensasse e così anche i denti trovati in Puglia nella Grotta del Cavallo di Uluzzo (provincia di Lecce) ne anticipano la presenza di 4.000 anni.
Un'altra ondata migratoria, attraverso l'Asia, avrebbe raggiunto l'America settentrionale attorno a 35/25.000 anni fa e poi 25/12.000 anni fa avrebbe colonizzato anche l'America centrale e meridionale.

(Clicca sull'immagine per ingrandire)
Ambiente e modo di vita
Questi ominidi erano in grado di adattarsi alle diverse condizioni ambientali - dal freddo artico alle aree desertiche -, anche quelle più difficili del periodo glaciale, entrando in competizione con i grandi predatori dell'Eurasia, sfruttando vegetazione e animali disponibili nelle regioni che colonizzavano, per questo sono riusciti a occupare progressivamente tutti i continenti.
Caratteristico del periodo antico fu l'uomo di Cro-Magnon, che popolava le regioni occidentali dell'Europa. Egli non solo produceva e utilizzava strumenti di pietra, ma li specializzava anche e produceva una varietà di strumenti più piccoli, più complessi, raffinati, tra cui utensili per raschiare, forare e incidere; erano capaci di lavorare le pelli, usando appositi raschiatoi, e fabbricavano ami per la pesca, aghi per cucire, punte per frecce e arpioni.

Ornamenti personali e strumenti in osso rinvenuti nella grotta di Bacho Kiro (Bulgaria, a sinistra) e nella Grotte du Renne (Francia, a destra). I manufatti della grotta di Bacho Kiro sono attribuiti all'Homo sapiens e risalgono a circa 45.000 anni fa; i manufatti della Grotte du Renne, realizzati millenni più tardi, sono attribuiti ai Neandertal (© Rosen Spasov and Geoff Smith, License: CC-BY-SA 2.0)
Il gusto estetico dell'uomo si raffinò sempre più e l'arte assunse un ruolo determinante nel progresso dell'umanità. Le caverne dell'Europa occidentale sono state utilizzate a scopo religioso o cultuale, come indicano le numerose raffigurazioni in esse ritrovate.


 Trascorreva gran parte della giornata a raccogliere piante e cacciare o scovare animali. La dieta era più varia di quanto si pensasse in passato perché non era costituita dalla sola cacciagione, ma comprendeva una grande varietà di vegetali, un'anticipazione della "dieta mediterranea". Aveva il controllo del fuoco e costruiva capanne per ripararsi e svolgere diverse attività.
Trascorreva gran parte della giornata a raccogliere piante e cacciare o scovare animali. La dieta era più varia di quanto si pensasse in passato perché non era costituita dalla sola cacciagione, ma comprendeva una grande varietà di vegetali, un'anticipazione della "dieta mediterranea". Aveva il controllo del fuoco e costruiva capanne per ripararsi e svolgere diverse attività.

Caratteristiche

 Le forme arcaiche di Homo sapiens presentano crani con caratteristiche intermedie tra H. erectus gli uomini moderni. Le dimensioni del cervello sono di 1200 cc, i denti sono meno robusti di H. erectus, il toro sopraorbitale è ancora ben evidente e il mento sfuggente.
Le forme arcaiche di Homo sapiens presentano crani con caratteristiche intermedie tra H. erectus gli uomini moderni. Le dimensioni del cervello sono di 1200 cc, i denti sono meno robusti di H. erectus, il toro sopraorbitale è ancora ben evidente e il mento sfuggente.
Nelle forme più evolute il cranio è nettamente diverso da quello dei Neandertaliani e da H. heidelbergensis, con perdita di robustezza, sia craniale, sia postcraniale: lo scheletro facciale è molto meno sviluppato, non è più così proiettato in avanti, la mandibola rivela un mento osseo pronunciato ed è meno robusta, con incisivi e i molari più piccoli. Non esiste più un toro sopraorbitario continuo, ma solo un leggero rilievo; il frontale e l'occipitale sono più arrotondati; lo chignon occipitale manca o è molto ridotto; la volta cranica è più elevata e con pareti sottili, con fronte spaziosa, piatta e quasi verticale e il cranio è più corto, con capacità analoga, o addirittura minore, rispetto a quella dei Neandertaliani (mediamente 1300 cc); il naso ha l'apertura nasale più stretta.
La discesa della laringe ha consentito lo sviluppo di un linguaggio articolato.
Lo scheletro è meno robusto di H. neanderthalensis e delle altre specie presenti in Eurasia e il bacino è diventato leggermente più stretto ai lati, ma più espanso e avvolgente verso l'avanti, aumentando in tal modo il canale del parto, una modifica richiesta dal vistoso aumento del nostro encefalo.
Implicazioni evolutive
Abbiamo già affrontato in precedenza il tema dell'origine dell'Homo sapiens. Riprendiamo qui alcuni concetti essenziali.
L'ipotesi prevalente è quella dell'origine unica africana (Out of Africa), intorno a 200/300.000 anni fa, a partire da una popolazione di H. ergaster che non era migrato in Asia, o da H. antecessor, o ancora da H. heidelbergensis/H. rodesiensis anch'essi rimasti in Africa.
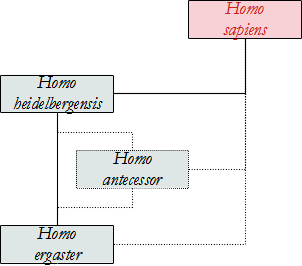
A questa teoria sono state proposte tre varianti: l'ipotesi del multiregionalismo africano, secondo la quale non ci sarebbe un unico luogo di origine in Africa, ma il moderno H. sapiens sarebbe emerso dalla confluenza di diverse popolazioni presenti sul continente; l'origine africana recente con ibridazione; il modello di assimilazione, a cui si rimanda per i dettagli.
Quando H. sapiens è migrato dall'Africa in Asia e in Europa ha sostituito completamente (o assimilato) le forme locali preesistenti di H. erectus, H. heidelbergensis, H. neanderthalensis e H. denisovensis, con le modalità descritte nei singoli Ominini.
Secondo il vecchio modello multiregionale, invece, H. ergaster, uscito originariamente dall'Africa circa due milioni di anni fa, avrebbe sviluppato negli altri continenti linee evolutive diverse le quali, mantenendo un continuo scambio genico, hanno dato origine in modo indipendente a H. sapiens nelle diverse etnie.
Una terza teoria, riproposta anche recentemente, è quella dell'origine euroasiatica (Out of Eurasia), seconda la quale H. erectus asiatico sarebbe ritornato in Africa dove avrebbe dato origine a H. sapiens.
Per quanto riguarda le possibili ibridazioni, ne abbiamo discusso affrontando i precedenti Ominini.