
In questa prima fase la luce scompone 12 molecole di H2O e libera 6 molecole di O2, mentre l'energia è accumulata in 12NADPH e 18ATP che saranno utilizzati nella fase oscura.
L'equazione complessiva della fase luminosa è la seguente.
![]()
Seguiamo nello schema i vari momenti della fase luminosa.

Innanzitutto la luce provoca la fotolisi di una molecola H2O presente nel lume del tilacoide, con liberazione di ½O2, 2H+ e 2e-. L'ossigeno prodotto, 6 molecole biatomiche in totale, è liberato nell'atmosfera.
![]()
Contemporaneamente, la luce colpisce il fotosistema II (P680) eccitando 2 elettroni che si portano verso il fotosistema I il quale, a sua volta, presenta un "buco" elettronico in quanto la luce ha scalzato una coppia di elettroni. Il buco di elettroni del fotosistema I, invece, è riempito con la coppia di elettroni provenienti dall'acqua.
La discesa spontanea degli elettroni verso il fotosistema I non avviene direttamente ma essi sono trasferiti dall'accettore primario (feofitina, una porfirina uguale alla clorofilla ma senza Mg) alla catena di trasporto degli elettroni presente nella membrana dei tilacoidi, costituita: plastochinone A (PQ), plastochinone B - complesso del citocromo b6f - plastocianina (PC), una proteina contenente Cu2+ che si riduce a Cu+.
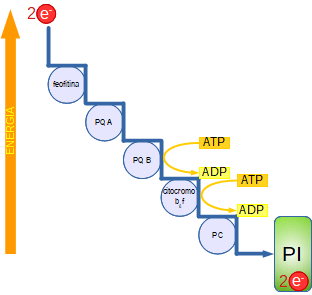
Poiché gli elettroni passano da un livello energetico alto a uno più basso, e quindi favorito termodinamicamente, si libera gradualmente energia che attiva la pompa protonica (complesso del citocromo b6f) (vedi la pagina iniziale) la quale, contro il gradiente di concentrazione, trasferisce gli ioni H+ (protoni) dallo stroma all'interno dei tilacoidi dove già sono abbondanti perché sono presenti anche quelli provenienti dalla fotolisi dell'acqua. Si genera così una pressione chemiosmotica.
I protoni tendono a ritornare nello stroma seguendo il gradiente di concentrazione ma, essendo la membrana impermeabile agli ioni, sono costretti a passare attraverso il canale del grande complesso enzimatico ATP-sintasi. Questo spostamento libera energia che viene impiegata per la sintesi di 2ATP (fotofosforilazione).
Gli elettroni del fotosistema I passano attraverso una seconda catena di trasporto: dall'accettore primario (fitochinone, una clorofilla modificata A0) a fitochinone A1- proteina ferro-zolfo (FeS) - ferredossina (Fd), una proteina contenente zolfo e Fe3+ che, con l'intervento di una flavoproteina (ferredossina NAPD+ reduttasi FNR) si riduce a Fe2+ e trasferisce gli elettroni al NADP+ insieme ai 2H+ prodotti nella fotolisi dell'acqua per formare NADPH (12 in totale). Uno solo degli ioni è però trasportato dal coenzima mentre l'altro rimane libero.

Poiché gli elettroni, da un composto iniziale fluiscono a un composto finale passando attraverso trasportatori intermedi, si parla di fotofosforilazione non ciclica.

Negli organismi che hanno solo il fotosistema I, o in base alle esigenze della cellula, gli elettroni non vanno al NADP+ (e quindi non si forma NADPH) ma dalla ferredossina ritornano al complesso del complesso del citocromo b6f, poi alla plastocianina e nuovamente convogliati al P700 (fotofosforilazione ciclica). Il pompaggio di H+ del citocromo genera potenziale chemiosmotico per la sintesi di ATP in quantità maggiore rispetto alla via non ciclica perché non c'è la spesa energetica per la produzione di glucosio.

Alla fine della fase luminosa abbiamo quindi molecole che hanno immagazzinato l'energia solare: 12NADPH e 18ATP pronte per essere utilizzate nella fase successiva, mentre l'O2 esce dagli stomi della foglia.